To assess the genetic variation of different SARS
CoV-2 strains, the 2019 Novel Coronavirus Resource
of China National Center for Bioinformation aligned
77,801 genome sequences of SARS-CoV-2 detected glob-
ally and identified a total of 15,018 mutations, including
14,824 single-nucleotide polymorphisms (BIGD)31.
In the S protein, four amino acid alterations, V483A,
L4551, F456V and G476S, are located near the binding
interface in the RBD, but their effects on binding to the
host receptor are unknown. The alteration D614G in
the S1 subunit was found far more frequently than other
S variant sites, and it is the marker of a major subclade of
SARS-CoV-2 (clade G). Since March 2020, SARS-CoV-2
variants with G614 in the S protein have replaced the
original D614 variants and become the dominant form
circulating globally. Compared with the D614 variant,
higher viral loads were found in patients infected with
the G614 variant, but clinical data suggested no signif-
icant link between the D614G alteration and disease
severity32. Pseudotyped viruses carrying the S protein
with G614 generated higher infectious titres than viruses
carrying the S protein with D614, suggesting the altera-
tion may have increased the infectivity of SARS-CoV-2
(REF.32). However, the results of in vitro experiments based
on pseudovirus, models may not exactly reflect natural
infection. This preliminary finding should be validated
by more studies using wild-type SARS-CoV-2 variants to
infect different target cells and animal models. Whether
this amino acid change enhanced virus transmissibil-
ity is also to be determined. Another marker mutation
for SARS-CoV-2 evolution is the single-nucleotide
appeared asymptomatic45. Another serological study
detected SARS-CoV-2 neutralizing antibodies in cat
serum samples collected in Wuhan after the COVID-19
outbreak, providing evidence for SARS-CoV-2 infection
in cat populations in Wuhan, although the potential
of SARS-CoV-2 transmission from cats to humans is
currently uncertain46.
Receptor use and pathogenesis
SARS-CoV-2 uses the same receptor as SARS-CoV,
angiotensin-converting enzyme 2 (ACE2) 11,47. Besides
human ACE2 (hACE2), SARS-CoV-2 also recognizes
ACE2 from pig, ferret, rhesus monkey, civet, cat, pan-
golin, rabbit and dog11,43,48,49. The broad receptor usage
of SARS-CoV-2 implies that it may have a wide host
range, and the varied efficiency of ACE2 usage in differ-
ent animals may indicate their different susceptibilities
to SARS-CoV-2 infection. The S1 subunit of a corona
virus is further divided into two functional domains,
an N-terminal domain and a C-terminal domain.
Structural and biochemical analyses identified a
211 amino acid region (amino acids 319–529) at the S1
C-terminal domain of SARS-CoV-2 as the RBD, which
has a key role in virus entry and is the target of neu
tralizing antibodies50,51 (FIG. 3a). The RBM mediates con-
tact with the ACE2 receptor (amino acids 437–507 of
SARS-CoV-2 S protein), and this region in SARS-CoV-2
differs from that in SARS-CoV in the five residues crit
INTRODUCTION
Over the past 2 decades, coronaviruses (CoVs)
have been associated with significant disease
outbreaks in East Asia and the Middle East. The
severe acute respiratory syndrome (SARS) and the
Middle East respiratory syndrome (MERS) began to
emerge in 2002 and 2012, respectively. Recently, a
novel coronavirus, severe acute respiratory
syndrome coronavirus 2 (SARS-CoV-2), causing
coronavirus disease 2019 (COVID-19), emerged in
late 2019, and it has posed a global health threat,
causing an ongoing pandemic in many countries and
territories (1).
Health workers worldwide are currently making
efforts to control further disease outbreaks caused by
the novel CoV (originally named 2019-nCoV),
which was first identified in Wuhan City, Hubei
Province, China, on 12 December 2019. On 11
February 2020, the World Health Organization
(WHO) announced the official designation for the
current CoV-associated disease to be COVID-19,
caused by SARS-CoV-2. The primary cluster of
patients was found to be connected with the Huanan
South China Seafood Market in Wuhan (2). COVs
belong to the family Coronaviridae (subfamily
Coronavirinae), the members of which infect a broad
range of hosts, producing symptoms and diseases
ranging from the common cold to severe and
ultimately fatal illnesses, such as SARS, MERS, and,
presently, COVID-19. SARS-CoV-2 is considered
one of the seven members of the CoV family that
infect humans (3), and it belongs to the same lineage
of CoVs that causes SARS; however, this novel virus
is genetically distinct. Until 2020, six CoVs were
known to infect humans, including human CoV 229E
(HCOV-229E), HCOV-NL63, HCOV-OC43, HCOV-
HKUL, SARS-CoV, and MERS-CoV. Although
SARS-CoV and MERS-CoV have resulted in
outbreaks with high mortality, others remain
associated with mild upper-respiratory-tract illnesses
(4).
Newly evolved CoVs pose a high threat to global
public health. The current emergence of COVID-19
is the third CoV outbreak in humans over the past 2
decades (5). It is no coincidence that Fan et al.
predicted potential SARS- or MERS-like COV
outbreaks in China following pathogen transmission
from bats (6). COVID-19 emerged in China and
spread rapidly throughout the country and,
subsequently, to other countries. Due to the severity
of this outbreak and the potential of spreading on an
international scale, the WHO declared a global
health emergency on 31 January 2020: subsequently
health emergency on 31 January 2020; subsequently,
on 11 March 2020, they declared it a pandemic
situation. At present, we are not in a position to
effectively treat COVID-19, since neither approved
vaccines nor specific antiviral drugs for treating
human CoV infections are available (7–9). Most
nations are currently making efforts to prevent the
further spreading of this potentially deadly virus by
implementing preventive and control strategies.
In domestic animals, infections with CoVs are
associated with a broad spectrum of pathological
conditions. Apart from infectious bronchitis virus,
canine respiratory CoV, and mouse hepatitis virus,
CoVs are predominantly associated with
gastrointestinal diseases (10). The emergence of
novel CoVs may have become possible because of
multiple CoVs being maintained in their natural host,
which could have favored the probability of genetic
recombination (10). High genetic diversity and the
ability to infect multiple host species are a result of
high-frequency mutations in CoVs, which occur due
to the instability of RNA-dependent RNA
polymerases along with higher rates of homologous
RNA recombination (10, 11). Identifying the origin
of SARS-CoV-2 and the pathogen’s evolution will be
helpful for disease surveillance (12), development of
new targeted drugs, and prevention of further
epidemics (13). The most common symptoms
associated with COVID-19 are fever, cough,
dyspnea, expectoration, headache, and myalgia or
fatigue.
In contrast, less common signs at the time of
hospital admission include diarrhea, hemoptysis, and
shortness of breath (14). Recently, individuals with
asymptomatic infections were also suspected of
transmitting infections, which further adds to the
complexity of disease transmission dynamics in
COVID-19 infections (1). Such efficient responses
require in-depth knowledge regarding the virus,
which currently is a novel agent; consequently,
further studies are required.
Comparing the genome of SARS-CoV-2 with that
of the closely related SARS/SARS-like COV
revealed that the sequence coding for the spike
protein, with a total length of 1,273 amino acids,
showed 27 amino acid substitutions. Six of these
substitutions are in the region of the receptor-binding
domain (RBD), and another six substitutions are in
the underpinning subdomain (SD) (16). Phylogenetic
analyses have revealed that SARS-CoV-2 is closely
related (88% similarity) to two SARS-like COVs
derived from bat SARS-like COVs (bat-SL
CoVZC45 and bat-SL-COVZXC21) (Fig. 1).
Furthermore, SARS-CoV-2 is genetically distinct
from SARS-CoV (79% similarity) and MERS-CoV
(nearly 50%) (17). COVID-19 is associated with
afflictions of the lungs in all cases and generated
characteristic chest computer tomography findings,
such as the presence of multiple lesions in lung lobes
that appear as dense, ground-glass opaque structures
that occasionally coexist with consolidation shadows
(18).
Some therapeutic options for treating COVID-19
showed efficacy in in vitro studies; however, to date,
these treatments have not undergone any randomized
animal or human clinical trials, which limit their
practical applicability in the current pandemic (7, 9,
19-21).
The present comprehensive review describes the
various features of SARS-CoV-2/COVID-19 causing
the current disease outbreaks and advances in
diagnosis and developing vaccines and therapeutics.
It also provides a brief comparison with the earlier
SARS and MERS COVs, the veterinary perspective
of CoVs and this emerging novel pathogen, and an
evaluation of the zoonotic potential of similar CoVs
to provide feasible One Health strategies for the
management of this fatal virus (22–367).
THE VIRUS (SARS-CoV-2)
Coronaviruses are positive-sense RNA viruses
having an extensive and promiscuous range of
natural hosts and affect multiple systems (23, 24).
Coronaviruses can cause clinical diseases in humans
that may extend from the common cold to more
severe respiratory diseases like SARS and MERS
(17, 279). The recently emerging SARS-CoV-2 has
wrought havoc in China and caused a pandemic
situation in the worldwide nonulation leading to
wrought havoc in China and caused a pandemic
situation in the worldwide population, leading to
disease outbreaks that have not been controlled to
date, although extensive efforts are being put in
place to counter this virus (25). This virus has been
proposed to be designated/named severe acute
respiratory syndrome coronavirus 2 (SARS-CoV-2)
by the International Committee on Taxonomy of
Viruses (ICTV), which determined the virus belongs
to the Severe acute respiratory syndrome-related
coronavirus category and found this virus is related
to SARS-CoVs (26). SARS-CoV-2 is a member of
the order Nidovirales, family Coronaviridae,
subfamily Orthocoronavirinae, which is subdivided
into four genera, viz., Alphacoronavirus,
Betacoronavirus, Gammacoronavirus, and
Deltacoronavirus (3, 27). The genera
Alphacoronavirus and Betacoronavirus originate
from bats, while Gammacoronavirus and
Deltacoronavirus have evolved from bird and swine
gene pools (24, 28, 29, 275).
Coronaviruses possess an unsegmented, single-
stranded, positive-sense RNA genome of around 30
kb, enclosed by a 5'-cap and 3'-poly(A) tail (30). The
genome of SARS-CoV-2 is 29,891 bp long, with a
G+C content of 38% (31). These viruses are
encircled with an envelope containing viral
encircled with an envelope containing viral
nucleocapsid. The nucleocapsids in CoVs are
arranged in helical symmetry, which reflects an
atypical attribute in positive-sense RNA viruses (30).
The electron micrographs of SARS-CoV-2 revealed
a diverging spherical outline with some degree of
pleomorphism, virion diameters varying from 60 to
140 nm, and distinct spikes of 9 to 12 nm, giving the
virus the appearance of a solar corona (3). The CoV
genome is arranged linearly as 5'-leader-UTR-
replicase-structural genes (S-E-M-N)-3' UTR-
poly(A) (32). Accessory genes, such as 3a/b, 4a/b,
and the hemagglutinin-esterase gene (HE), are also
seen intermingled with the structural genes (30).
SARS-CoV-2 has also been found to be arranged
similarly and encodes several accessory proteins,
although it lacks the HE, which is characteristic of
some betacoronaviruses (31). The positive-sense
genome of CoVs serves as the mRNA and is
translated to polyprotein la/lab (ppla/lab) (33). A
replication-transcription complex (RTC) is formed in
double-membrane vesicles (DMVs) by nonstructural
proteins (nsps), encoded by the polyprotein gene
(34). Subsequently, the RTC synthesizes a nested set
of subgenomic RNAs (sgRNAs) via discontinuous
transcription (35).
Based on molecular characterization, SARSC
oV-2 is considered a new Betacoronavirus
belonging to the subgenus Sarbecovirus (3). A few
other critical zoonotic viruses (MERS-related CoV
and SARS-related CoV) belong to the same genus.
However, SARS-CoV-2 was identified as a distinct
virus based on the percent identity with other
Betacoronavirus; conserved open reading frame 1a/b
(ORFla/b) is below 90% identity (3). An overall
80% nucleotide identity was observed between
SARS-CoV-2 and the original SARS-CoV, along
with 89% identity with ZC45 and ZXC21 SARS
related CoVs of bats (2, 31, 36). In addition, 82%
identity has been observed between SARS-CoV-2
and human SARS-CoV Tor2 and human SARS-CoV
BJ01 2003 (31). A sequence identity of only 51.8%
was observed between MERS-related CoV and the
recently emerged SARS-CoV-2 (37). Phylogenetic
analysis of the structural genes also revealed that
SARS-CoV-2 is closer to bat SARS-related CoV.
Therefore, SARS-CoV-2 might have originated from
bats, while other amplifier hosts might have played a
role in disease transmission to humans (31). Of note,
the other two zoonotic CoVs (MERS-related CoV
and SARS-related CoV) also originated from bats
(38, 39). Nevertheless, for SARS and MERS, civet
(30, 59). Nevertheless, TOT SARS and MERS, Civet
cat and camels, respectively, act as amplifier hosts
(40, 41).
Coronavirus genomes and subgenomes encode
six ORFs (31). The majority of the 5' end is occupied
by ORF1a/b, which produces 16 nsps. The two
polyproteins, ppla and pplab, are initially produced
from ORF1a/b by a -1 frameshift between ORFla
and ORF16 (32). The virus-encoded proteases cleave
polyproteins into individual nsps (main protease
[Mpro), chymotrypsin-like protease [3CL pro], and
papain-like proteases [PLPs]) (42). SARS-CoV-2
also encodes these nsps, and their functions have
been elucidated recently (31). Remarkably, a
difference between SARS-CoV-2 and other CoVs is
the identification of a novel short putative protein
within the ORF3 band, a secreted protein with an
alpha helix and beta-sheet with six strands encoded
by ORF8 (31).
Coronaviruses encode four major structural
proteins, namely, spike (S), membrane (M), envelope
(E), and nucleocapsid (N), which are described in
detail below.
S Glycoprotein
Coronavirus S protein is a large, multifunctional
class I viral transmembrane protein. The size of this
Coronavirus S protein is a large, multifunctional
class I viral transmembrane protein. The size of this
abundant S protein varies from 1,160 amino acids
(IBV, infectious bronchitis virus, in poultry) to 1,400
amino acids (FCOV, feline coronavirus) (43). It lies
in a trimer on the virion surface, giving the virion a
corona or crown-like appearance. Functionally it is
required for the entry of the infectious virion
particles into the cell through interaction with
various host cellular receptors (44).
Furthermore, it acts as a critical factor for tissue
tropism and the determination of host range (45).
Notably, S protein is one of the vital
immunodominant proteins of COVs capable of
inducing host immune responses (45). The
ectodomains in all CoVs S proteins have similar
domain organizations, divided into two subunits, S1
and S2 (43). The first one, S1, helps in host receptor
binding, while the second one, S2, accounts for
fusion. The former (S1) is further divided into two
subdomains, namely, the N-terminal domain (NTD)
and C-terminal domain (CTD). Both of these
subdomains act as receptor-binding domains,
interacting efficiently with various host receptors
(45). The S1 CTD contains the receptor-binding
motif (RBM). In each coronavirus spike protein, the
trimeric S1 locates itself on top of the trimeric S2
trimeric Si locates itself on top of the trimeric S2
stalk (45). Recently, structural analyses of the S
proteins of COVID-19 have revealed 27 amino acid
substitutions within a 1,273-amino-acid stretch (16).
Six substitutions are located in the RBD (amino
acids 357 to 528), while four substitutions are in the
RBM at the CTD of the S1 domain (16). Of note, no
amino acid change is seen in the RBM, which binds
directly to the angiotensin-converting enzyme-2
(ACE2) receptor in SARS-CoV (16, 46). At present,
the main emphasis is knowing how many differences
would be required to change the host tropism.
Sequence comparison revealed 17 nonsynonymous
changes between the early sequence of SARS-CoV-2
and the later isolates of SARS-CoV. The changes
were found scattered over the genome of the virus,
with nine substitutions in ORFlab, ORF8 (4
substitutions), the spike gene (3 substitutions), and
ORF7a (single substitution) (4). Notably, the same
nonsynonymous changes were found in a familial
cluster, indicating that the viral evolution happened
during person-to-person transmission (4, 47). Such
adaptive evolution events are frequent and constitute
a constantly ongoing process once the virus spreads
among new hosts (47). Even though no functional
changes occur in the virus associated with this
adaptive evolution, close monitoring of the viral
adaptive evolution, close monitoring of the viral
mutations that occur during subsequent human-to-
human transmission is warranted.
M Protein
The M protein is the most abundant viral protein
present in the virion particle, giving a definite shape
to the viral envelope (48). It binds to the
nucleocapsid and acts as a central organizer of
coronavirus assembly (49). Coronavirus M proteins
are highly diverse in amino acid contents but
maintain overall structural similarity within different
genera (50). The M protein has three transmembrane
domains, flanked by a short amino terminus outside
the virion and a long carboxy terminus inside the
virion (50). Overall, the viral scaffold is maintained
by M-M interaction. Of note, the M protein of
SARS-CoV-2 does not have an amino acid
substitution compared to that of SARS-CoV (16).
E Protein
The coronavirus E protein is the most enigmatic
and smallest of the major structural proteins (51). It
plays a multifunctional role in the pathogenesis,
assembly, and release of the virus (52). It is a small
integral membrane polypeptide that acts as a
viroporin (ion channel) (53). The inactivation or
absence of this protein is related to the altered
virulence of coronaviruses due to changes in
morphology and tropism (54). The E protein consists
of three domains, namely, a short hydrophilic amino
terminal, a large hydrophobic transmembrane
domain, and an efficient C-terminal domain (51).
The SARS-CoV-2 E protein reveals a similar amino
acid constitution without any substitution (16).
N Protein
The N protein of coronavirus is multipurpose.
Among several functions, it plays a role in complex
formation with the viral genome, facilitates M
protein interaction needed during virion assembly,
and enhances the transcription efficiency of the virus
(55, 56). It contains three highly conserved and
distinct domains, namely, an NTD, an RNA-binding
domain or a linker region (LKR), and a CTD (57).
The NTD binds with the 3' end of the viral genome,
perhaps via electrostatic interactions, and is highly
diverged both in length and sequence (58). The
charged LKR is serine and arginine rich and is also
known as the SR (serine and arginine) domain (59).
The LKR is capable of direct interaction with in vitro
RNA interaction and is responsible for cell signaling
(60, 61).It also modulates the antiviral response of
the host by working as an antagonist for interferon
N Protein
The N protein of coronavirus is multipurpose.
Among several functions, it plays a role in complex
formation with the viral genome, facilitates M
protein interaction needed during virion assembly,
and enhances the transcription efficiency of the virus
(55, 56). It contains three highly conserved and
distinct domains, namely, an NTD, an RNA-binding
domain or a linker region (LKR), and a CTD (57).
The NTD binds with the 3' end of the viral genome,
perhaps via electrostatic interactions, and is highly
diverged both in length and sequence (58). The
charged LKR is serine and arginine rich and is also
known as the SR (serine and arginine) domain (59).
The LKR is capable of direct interaction with in vitro
RNA interaction and is responsible for cell signaling
(60, 61). It also modulates the antiviral response of
the host by working as an antagonist for interferon
(IFN) and RNA interference (62). Compared to that
of SARS-CoV, the N protein of SARS-CoV-2
possess five amino acid mutations, where two are in
the intrinsically dispersed region (IDR; positions 25
and 26), one each in the NTD (position 103), LKR
(position 217), and CTD (position 334) (16).
nsps and Accessory Proteins
nsps and Accessory Proteins
Besides the important structural proteins, the
SARS-CoV-2 genome contains 15 nsps, nspl to
nsp10 and nsp12 to nsp16, and 8 accessory proteins
(3a, 3b, p6, 7a, 7b, 8b, 9b, and ORF14) (16). All
these proteins play a specific role in viral replication
(27). Unlike the accessory proteins of SARS-CoV,
SARS-CoV-2 does not contain 8a protein and has a
longer 8b and shorter 3b protein (16). The nsp7,
nsp13, envelope, matrix, and p6 and 8b accessory
proteins have not been detected with any amino acid
substitutions compared to the sequences of other
coronaviruses (16).
The virus structure of SARS-CoV-2 is depicted in
Fig. 2.

We assessed the nucleotide percent similarity
using the MegAlign software program, where the
similarity between the novel SARS-CoV-2 isolates
was in the range of 99.4% to 100%. Among the other
Serbecovirus CoV sequences, the novel SARS-CoV-
2 sequences revealed the highest similarity to bat-
SL-CoV, with nucleotide percent identity ranges
between 88.12 and 89.65%. Meanwhile, earlier
reported SARS-CoVs showed 70.6 to 74.9%
similarity to SARS-CoV-2 at the nucleotide level.
Further, the nucleotide percent similarity was 55.4%,
45.5% to 47.9%, 46.2% to 46.6%, and 45.0% to
46.3% to the other four subgenera, namely,
Hibecovirus, Nobecovirus, Merbecovirus, and
Embecovirus, respectively. The percent similarity
index of current outbreak isolates indicates a close
relationship between SARS-CoV-2 isolates and bat-
SL-CoV, indicating a common origin. However,
particular pieces of evidence based on further
complete genomic analysis of current isolates are
necessary to draw any conclusions, although it was
ascertained that the current novel SARS-CoV-2
isolates belong to the subgenus Sarbecovirus in the
diverse range of beta coronaviruses. Their possible
ancestor was hypothesized to be from bat CoV
strains, wherein bats might have played a crucial role
in harboring this class of viruses.
Splits Tree phylogeny analysis.
In the unrooted phylogenetic tree of different
betacoronaviruses based on the S protein, virus
sequences from different subgenera grouped into
separate clusters. SARS-CoV-2 sequences from
Wuhan and other countries exhibited a close
relationship and appeared in a single cluster (Fig. 1).
The CoVs from the subgenus Sarbecovirus appeared
jointly in SplitsTree and divided into three
subclusters, namely, SARS-CoV-2, bat-SARS-like-
CoV (bat-SL-CoV), and SARS-CoV (Fig. 1). In the
case of other subgenera, like Merbecovirus, all of the
sequences grouped in a single cluster, whereas in
Embecovirus, different species, comprised of canine
respiratory CoVs, bovine CoVs, equine CoVs, and
human CoV strain (OC43), grouped in a common
cluster. Isolates in the subgenera Nobecovorus and
Hibecovirus were found to be placed separately
away from other reported SARS-CoVs but shared a
bat origin.
CURRENT WORLDWIDE SCENARIO OF
SARS-CoV-2
This novel virus, SARS-CoV-2, comes under the
subgenus Sarbecovirus of the Orthocoronavirinae
subfamily and is entirely different from the viruses
responsible for MERS-CoV and SARS-CoV (3). The
newly emerged SARS-CoV-2 is a group 2B
coronavirus (2). The genome sequences of SARS-
CoV-2 obtained from patients share 79.5% sequence
similarity to the sequence of SARS-CoV (63).
As of 13 May 2020, a total of 4,170,424
confirmed cases of COVID-19 (with 287,399 deaths)
have been reported in more than 210 affected
countries worldwide (WHO Situation Report 114
Initially, the epicenter of the SARS-CoV-2
pandemic was China, which reported a significant
number of deaths associated with COVID-19, with
84,458 laboratory-confirmed cases and 4,644 deaths
as of 13 May 2020 (Fig. 4). As of 13 May 2020,
SARS-CoV-2 confirmed cases have been reported in
more than 210 countries apart from China (Fig. 3
and 4) (WHO Situation Report 114) (25, 64).
COVID-19 has been reported on all continents
except Antarctica. For many weeks, Italy was the
focus of concerns regarding the large number of
cases, with 221,216 cases and 30,911 deaths, but
now, the United States is the country with the largest
number of cases, 1,322,054, and 79,634 deaths.
Now, the United Kingdom has even more cases
(226,4671) and deaths (32,692) than Italy. A John
Hopkins University web platform has provided daily
updates on the basic epidemiology of the COVID-19
outbreak
COVID-19 has also been confirmed on a cruise
ship, named Diamond Princess, quarantined in
Japanese waters (Port of Yokohama), as well as on
other cruise ships around the world (239) (Fig. 3).
The significant events of the SARS-CoV-2/COVID-
19 virus outbreak occurring since 8 December 2019
are presented as a timeline in Fig. 5.

FIG 5 Timeline depicting the significant events that
occurred during the SARS-CoV-2/COVID-19 virus
outbreak. The timeline describes the significant events
during the current SARS-CoV-2 outbreak, from 8
December 2019 to 13 May 2020.
At the beginning, China experienced the majority
of the burden associated with COVID-19 in the form
of disease morbidity and mortality (65), but over
time the COVID-19 menace moved to Europe,
particularly Italy and Spain, and now the United
States has the highest number of confirmed cases
possible origin of SARS-CO V-2 and the first mode of
disease transmission are not yet identified (70).
Analysis of the initial cluster of infections suggests
that the infected individuals had a common exposure
point, a seafood market in Wuhan, Hubei Province,
China (Fig. 6). The restaurants of this market are
well-known for providing different types of wild
animals for human consumption (71). The Huanan
South China Seafood Market also sells live animals,
such as poultry, bats, snakes, and marmots (72). This
might be the point where zoonotic (animal-to-
human transmission occurred (71). Although
SARS-CoV-2 is alleged to have originated from an
animal host (zoonotic origin) with further human-to-
human transmission (Fig. 6), the likelihood of
foodborne transmission should be ruled out with
further investigations, since it is a latent possibility
(1). Additionally, other potential and expected routes
would be associated with transmission, as in other
respiratory viruses, by direct contact, such as shaking
contaminated hands, or by direct contact with
contaminated surfaces (Fig. 6). Still, whether blood
transfusion and organ transplantation (276), as well
as transplacental and perinatal routes, are possible
routes for SARS-CoV-2 transmission needs to be
determined (Fig. 6).
From experience with several outbreaks
associated with known emerging viruses, higher
pathogenicity of a virus is often associated with
lower transmissibility. Compared to emerging
viruses like Ebola virus, avian H7N9, SARS-CoV,
and MERS-CoV, SARS-CoV-2 has relatively lower
pathogenicity and moderate transmissibility (15).
The risk of death among individuals infected with
COVID-19 was calculated using the infection
fatality risk (IFR). The IFR was found to be in the
range of 0.3% to 0.6%, which is comparable to that
of a previous Asian influenza pandemic (1957 to
1958) (73, 277).
Notably, the reanalysis of the COVID-19
pandemic curve from the initial cluster of cases
pointed to considerable human-to-human
transmission. It is opined that the exposure history of
SARS-CoV-2 at the Wuhan seafood market
originated from human-to-human transmission rather
than animal-to-human transmission (74); however, in
light of the zoonotic spillover in COVID-19, is too
early to fully endorse this idea (1). Following the
initial infection, human-to-human transmission has
been observed with a preliminary reproduction
number (Ro) estimate of 1.4 to 2.5 (70, 75), and
recently it is estimated to be 2.24 to 3.58 (76). In
another study, the average reproductive number of
another study, the average reproductive number of
COVID-19 was found to be 3.28, which is
significantly higher than the initial WHO estimate of
1.4 to 2.5 (77). It is too early to obtain the exact Ro
value, since there is a possibility of bias due to
insufficient data. The higher Ro value is indicative of
the more significant potential of SARS-CoV-2
transmission in a susceptible population. This is not
the first time where the culinary practices of China
have been blamed for the origin of novel coronavirus
infection in humans. Previously, the animals present
in the live-animal market were identified to be the
intermediate hosts of the SARS outbreak in China
(78). Several wildlife species were found to harbor
potentially evolving coronavirus strains that can
overcome the species barrier (79). One of the main
principles of Chinese food culture is that live-
slaughtered animals are considered more nutritious
(5).
After 4 months of struggle that lasted from
December 2019 to March 2020, the COVID-19
situation now seems under control in China. The wet
animal markets have reopened, and people have
started buying bats, dogs, cats, birds, scorpions,
badgers, rabbits, pangolins (scaly anteaters), minks,
soup from palm civet, ostriches, hamsters, snapping
turtles, ducks, fish, Siamese crocodiles, and other
turtles, ducks, fish, Siamese crocodiles, and other
animal meats without any fear of COVID-19. The
Chinese government is encouraging people to feel
they can return to normalcy. However, this could be
a risk, as it has been mentioned in advisories that
people should avoid contact with live-dead animals
as much as possible, as SARS-CoV-2 has shown
zoonotic spillover. Additionally, we cannot rule out
the possibility of new mutations in the same virus
being closely related to contact with both animals
and humans at the market (284). In January 2020,
China imposed a temporary ban on the sale of live-
dead animals in wet markets. However, now
hundreds of such wet markets have been reopened
without optimizing standard food safety and
sanitation practices (286).
With China being the most populated country in
the world and due to its domestic and international
food exportation policies, the whole world is now
facing the menace of COVID-19, including China
itself. Wet markets of live-dead animals do not
maintain strict food hygienic practices. Fresh blood
splashes are present everywhere, on the floor and
tabletops, and such food customs could encourage
many pathogens to adapt, mutate, and jump the
species barrier. As a result, the whole world is
suffering from novel SARS-CoV-2, with more than
suffering from novel SARS-CoV-2, with more than
4,170,424 cases and 287,399 deaths across the globe.
There is an urgent need for a rational international
campaign against the unhealthy food practices of
China to encourage the sellers to increase hygienic
food practices or close the crude live-dead animal
wet markets. There is a need to modify food policies
at national and international levels to avoid further
life threats and economic consequences from any
emerging or reemerging pandemic due to close
animal-human interaction (285).
Even though individuals of all ages and sexes are
susceptible to COVID-19, older people with an
underlying chronic disease are more likely to
become severely infected (80). Recently, individuals
with asymptomatic infection were also found to act
as a source of infection to susceptible individuals
(81). Both the asymptomatic and symptomatic
patients secrete similar viral loads, which indicates
that the transmission capacity of asymptomatic or
minimally symptomatic patients is very high. Thus,
SARS-CoV-2 transmission can happen early in the
course of infection (82). Atypical clinical
manifestations have also been reported in COVID-19
in which the only reporting symptom was fatigue.
Such patients may lack respiratory signs, such as
fever, cough, and sputum (83). Hence, the clinicians
fever, cough, and sputum (83). Hence, the clinicians
must be on the look-out for the possible occurrence
of atypical clinical manifestations to avoid the
possibility of missed diagnosis. The early
transmission ability of SARS-CoV-2 was found to be
similar to or slightly higher than that of SARS-CoV,
reflecting that it could be controlled despite
moderate to high transmissibility (84).
Increasing reports of SARS-CoV-2 in sewage and
wastewater warrants the need for further
investigation due to the possibility of fecal-oral
transmission. SARS-CoV-2 present in environmental
compartments such as soil and water will finally end
up in the wastewater and sewage sludge of treatment
plants (328). Therefore, we have to reevaluate the
current wastewater and sewage sludge treatment
procedures and introduce advanced techniques that
are specific and effective against SARS-CoV-2.
Since there is active shedding of SARS-CoV-2 in the
stool, the prevalence of infections in a large
population can be studied using wastewater-based
epidemiology. Recently, reverse transcription-
quantitative PCR (RT-qPCR) was used to enumerate
the copies of SARS-CoV-2 RNA concentrated from
wastewater collected from a wastewater treatment
plant (327). The calculated viral RNA copy numbers
determine the number of infected individuals. The
route warrants the introduction of negative fecal viral
nucleic acid test results as one of the additional
discharge criteria in laboratory-confirmed cases of
COVID-19 (326).
The COVID-19 pandemic does not have any
novel factors, other than the genetically unique
pathogen and a further possible reservoir. The cause
and the likely future outcome are just repetitions of
our previous interactions with fatal coronaviruses.
The only difference is the time of occurrence and the
genetic distinctness of the pathogen involved.
Mutations on the RBD of CoVs facilitated their
capability of infecting newer hosts, thereby
expanding their reach to all corners of the world
(85). This is a potential threat to the health of both
animals and humans. Advanced studies using
Bayesian phylogeographic reconstruction identified
the most probable origin of SARS-CoV-2 as the bat
SARS-like coronavirus, circulating in the
Rhinolophus bat family (86).
Phylogenetic analysis of 10 whole-genome
sequences of SARS-CoV-2 showed that they are
related to two CoVs of bat origin, namely, bat-SL-
CoVZC45 and bat-SL-COVZXC21, which were
reported during 2018 in China (17). It was reported
that SARS-CoV-2 had been confirmed to use ACE2
as an entry receptor while exhibiting an RBD similar
as an entry receptor while exhibiting an RBD similar
to that of SARS-CoV (17, 87, 254, 255). Several
countries have provided recommendations to their
people traveling to China (88, 89). Compared to the
previous coronavirus outbreaks caused by SARS-
CoV and MERS-CoV, the efficiency of SARS-CoV-
2 human-to-human transmission was thought to be
less. This assumption was based on the finding that
health workers were affected less than they were in
previous outbreaks of fatal coronaviruses (2).
Superspreading events are considered the main
culprit for the extensive transmission of SARS and
MERS (90, 91). Almost half of the MERS-CoV
cases reported in Saudi Arabia are of secondary
origin that occurred through contact with infected
asymptomatic or symptomatic individuals through
human-to-human transmission (92). The occurrence
of superspreading events in the COVID-19 outbreak
cannot be ruled out until its possibility is evaluated.
Like SARS and MERS, COVID-19 can also infect
the lower respiratory tract, with milder symptoms
(27). The basic reproduction number of COVID-19
has been found to be in the range of 2.8 to 3.3 based
on real-time reports and 3.2 to 3.9 based on predicted
infected cases (84).
Coronaviruses in Humans—SARS, MERS,
and COVID-19
Coronavirus infection in humans is commonly
associated with mild to severe respiratory diseases,
with high fever, severe inflammation, cough, and
internal organ dysfunction that can even lead to
death (92). Most of the identified coronaviruses
cause the common cold in humans. However, this
changed when SARS-CoV was identified, paving the
way for severe forms of the disease in humans (22).
Our previous experience with the outbreaks of other
coronaviruses, like SARS and MERS, suggests that
the mode of transmission in COVID-19 as mainly
human-to-human transmission via direct contact,
droplets, and fomites (25). Recent studies have
demonstrated that the virus could remain viable for
hours in aerosols and up to days on surfaces; thus,
aerosol and fomite contamination could play potent
roles in the transmission of SARS-CoV-2 (257).
The immune response against coronavirus is vital
to control and get rid of the infection. However,
maladjusted immune responses may contribute to the
immunopathology of the disease, resulting in
impairment of pulmonary gas exchange.
Understanding the interaction between CoVs and
host innate immune systems could enlighten our
understanding of the lung inflammation associated
with this infection (24).
SARS is a viral respiratory disease caused by a
formerly unrecognized animal CoV that originated
from the wet markets in southern China after
adapting to the human host, thereby enabling
transmission between humans (90). The SARS
outbreak reported in 2002 to 2003 had 8,098
confirmed cases with 774 total deaths (9.6%) (93).
The outbreak severely affected the Asia Pacific
region, especially mainland China (94). Even though
the case fatality rate (CFR) of SARS-CoV-2
(COVID-19) is lower than that of SARS-CoV, there
exists a severe concern linked to this outbreak due to its epidemiological similarity to influenza viruses
(95, 279). This can fail the public health system,
resulting in a pandemic (96).
MERS is another respiratory disease that was
first reported in Saudi Arabia during the year 2012.
The disease was found to have a CFR of around 35%
(97). The analysis of available data sets suggests that
the incubation period of SARS-CoV-2, SARS-CoV,
and MERS-CoV is in almost the same range. The
longest predicted incubation time of SARS-CoV-2 is
14 days. Hence, suspected individuals are isolated
for 14 days to avoid the risk of further spread (98).
Even though a high similarity has been reported
Even though a high similarity has been reported
between the genome sequence of the new
coronavirus (SARS-CoV-2) and SARS-like CoVs,
the comparative analysis recognized a furin-like
cleavage site in the SARS-CoV-2 S protein that is
missing from other SARS-like COVs (99). The furin-
like cleavage site is expected to play a role in the life
cycle of the virus and disease pathogenicity and
might even act as a therapeutic target for furin
inhibitors. The highly contagious nature of SARS-
CoV-2 compared to that of its predecessors might be
the result of a stabilizing mutation that occurred in
the endosome-associated-protein-like domain of
nsp2 protein.
Similarly, the destabilizing mutation near the
phosphatase domain of nsp3 proteins in SARS-CoV-
2 could indicate a potential mechanism that
differentiates it from other CoVs (100). Even though
the CFR reported for COVID-19 is meager
compared to those of the previous SARS and MERS
outbreaks, it has caused more deaths than SARS and
MERS combined (101). Possibly related to the viral
pathogenesis is the recent finding of an 832
nucleotide (nt) deletion in ORF8, which appears to
reduce the replicative fitness of the virus and leads to
attenuated phenotypes of SARS-CoV-2 (256).
Coronavirus is the most prominent example of a
Coronavirus is the most prominent example of a
virus that has crossed the species barrier twice from
wild animals to humans during SARS and MERS
outbreaks (79, 102). The possibility of crossing the
species barrier for the third time has also been
suspected in the case of SARS-CoV-2 (COVID-19).
Bats are recognized as a possible natural reservoir
host of both SARS-CoV and MERS-CoV infection.
In contrast, the possible intermediary host is the
palm civet for SARS-CoV and the dromedary camel
for MERS-CoV infection (102). Bats are considered
the ancestral hosts for both SARS and MERS (103).
Bats are also considered the reservoir host of human
coronaviruses like HC0V-229E and HCOV-NL63
(104). In the case of COVID-19, there are two
possibilities for primary transmission: it can be
transmitted either through intermediate hosts, similar
to that of SARS and MERS, or directly from bats
(103). The emergence paradigm put forward in the
SARS outbreak suggests that SARS-CoV originated
from bats (reservoir host) and later jumped to civets
(intermediate host) and incorporated changes within
the receptor-binding domain (RBD) to improve
binding to civet ACE2. This civet-adapted virus,
during their subsequent exposure to humans at live
markets, promoted further adaptations that resulted
in the epidemic strain (104). Transmission can also
markets, promoted Turther adaptations that resulted
in the epidemic strain (104). Transmission can also
occur directly from the reservoir host to humans
without RBD adaptations. The bat coronavirus that is
currently in circulation maintains specific “poised”
spike proteins that facilitate human infection without
the requirement of any mutations or adaptations
(105). Altogether, different species of bats carry a
massive number of coronaviruses around the world
(106).
The high plasticity in receptor usage, along with
the feasibility of adaptive mutation and
recombination, may result in frequent interspecies
transmission of coronavirus from bats to animals and
humans (106). The pathogenesis of most bat
coronaviruses is unknown, as most of these viruses
are not isolated and studied (4). Hedgehog
coronavirus HKU31, a Betacoronavirus, has been
identified from amur hedgehogs in China. Studies
show that hedgehogs are the reservoir of
Betacoronavirus, and there is evidence of
recombination (107).
The current scientific evidence available on
MERS infection suggests that the significant
reservoir host, as well as the animal source of MERS
infection in humans, is the dromedary camels (97).
The infected dromedary camels may not show any
visible signs of infection. making it challenging to
visible signs of infection, making it challenging to
identify animals actively excreting MERS-CoV that
has the potential to infect humans. However, they
may shed MERS-CoV through milk, urine, feces,
and nasal and eye discharge and can also be found in
the raw organs (108). In a study conducted to
evaluate the susceptibility of animal species to
MERS-CoV infection, llamas and pigs were found to
be susceptible, indicating the possibility of MERS-
CoV circulation in animal species other than
dromedary camels (109).
Following the outbreak of SARS in China,
SARS-CoV-like viruses were isolated from
Himalayan palm civets (Paguma larvata) and
raccoon dogs (Nyctereutes procyonoides) found in a
live-animal market in Guangdong, China. The
animal isolates obtained from the live-animal market
retained a 29-nucleotide sequence that was not
present in most of the human isolates (78). These
findings were critical in identifying the possibility of
interspecies transmission in SARS-CoV. The higher
diversity and prevalence of bat coronaviruses in this
region compared to those in previous reports indicate
a host/pathogen coevolution. SARS-like
coronaviruses also have been found circulating in the
Chinese horseshoe bat (Rhinolophus sinicus)
populations. The in vitro and in vivo studies carried
populations. The in vitro and in vivo studies carried
out on the isolated virus confirmed that there is a
potential risk for the reemergence of SARS-CoV
infection from the viruses that are currently
circulating in the bat population (105).
CLINICAL PATHOLOGY OF SARS-CoV-2
(COVID-19)
The disease caused by SARS-CoV-2 is also
named severe specific contagious pneumonia
(SSCP), Wuhan pneumonia, and, recently, COVID-
19 (110). Compared to SARS-CoV, SARS-CoV-2
has less severe pathogenesis but has superior
transmission capability, as evidenced by the rapidly
increasing number of COVID-19 cases (111). The
incubation period of SARS-CoV-2 in familial
clusters was found to be 3 to 6 days (112). The mean
incubation period of COVID-19 was found to be 6.4
days, ranging from 2.1 to 11.1 days (113). Among an
early affected group of 425 patients, 59 years was the
median age, of which more males were affected
(114). Similar to SARS and MERS, the severity of
this nCoV is high in age groups above 50 years (2,
115). Symptoms of COVID-19 include fever, cough,
myalgia or fatigue, and, less commonly, headache,
hemoptysis, and diarrhea (116, 282). Compared to
the SARS-CoV-2-infected patients in Wuhan during
the initial stages of the outbreak, only mild
symptoms were noticed in those patients that are
infected by human-to-human transmission (14).
The initial trends suggested that the mortality
associated with COVID-19 was less than that of
previous outbreaks of SARS (101). The updates
obtained from countries like China, Japan, Thailand,
and South Korea indicated that the COVID-19
patients had relatively mild manifestations compared
to those with SARS and MERS (4). Regardless of
the coronavirus type, immune cells, like mast cells,
that are present in the submucosa of the respiratory
tract and nasal cavity are considered the primary
barrier against this virus (92). Advanced in-depth
analysis of the genome has identified 380 amino acid
substitutions between the amino acid sequences of
SARS-CoV-2 and the SARS/SARS-like
coronaviruses. These differences in the amino acid
sequences might have contributed to the difference
in the pathogenic divergence of SARS-CoV-2 (16).
Further research is required to evaluate the possible
differences in tropism, pathogenesis, and
transmission of this novel agent associated with this
change in the amino acid sequence. With the current
outbreak of COVID-19, there is an expectancy of a
significant increase in the number of published
studies about this emerging coronavirus, as occurred
with SARS and MERS (117).
SARS-CoV-2 invades the lung parenchyma,
resulting in severe interstitial inflammation of the
lungs. This is evident on computed tomography (CT)
images as ground-glass opacity in the lungs. This
lesion initially involves a single lobe but later
expands to multiple lung lobes (118). The
histological assessment of lung biopsy samples
obtained from COVID-19-infected patients revealed
diffuse alveolar damage, cellular fibromyxoid
exudates, hyaline membrane formation, and
desquamation of pneumocytes, indicative of acute
respiratory distress syndrome (119). It was also
found that the SARS-CoV-2-infected patients often
have lymphocytopenia with or without leukocyte
abnormalities. The degree of lymphocytopenia gives
an idea about disease prognosis, as it is found to be
positively correlated with disease severity (118).
Pregnant women are considered to have a higher risk
of getting infected by COVID-19. The coronaviruses
can cause adverse outcomes for the fetus, such as
intrauterine growth restriction, spontaneous abortion,
preterm delivery, and perinatal death.
Nevertheless, the possibility of intrauterine
maternal-fetal transmission (vertical transmission) of
CoVs is low and was not seen during either the
SARS- or MERS-CoV outbreak (120). However,
SARS- or MERS-CoV outbreak (120). However,
there has been concern regarding the impact of
SARS-CoV-2/COVID-19 on pregnancy. Researchers
have mentioned the probability of in utero
transmission of novel SARS-CoV-2 from COVID-
19-infected mothers to their neonates in China based
upon the rise in IgM and IgG antibody levels and
cytokine values in the blood obtained from newborn
infants immediately postbirth; however, RT-PCR
failed to confirm the presence of SARS-CoV-2
genetic material in the infants (283). Recent studies
show that at least in some cases, preterm delivery
and its consequences are associated with the virus.
Nonetheless, some cases have raised doubts for the
likelihood of vertical transmission (240–243).
COVID-19 infection was associated with
pneumonia, and some developed acute respiratory
distress syndrome (ARDS). The blood biochemistry
indexes, such as albumin, lactate dehydrogenase, C-
reactive protein, lymphocytes (percent), and
neutrophils (percent) give an idea about the disease
severity in COVID-19 infection (121). During
COVID-19, patients may present leukocytosis,
leukopenia with lymphopenia (244),
hypoalbuminemia, and an increase of lactate
dehydrogenase, aspartate transaminase, alanine
aminotransferase, bilirubin, and, especially, D-dimer
aminotransferase, bilirubin, and, especially, D-dimer
(244). Middle-aged and elderly patients with primary
chronic diseases, especially high blood pressure and
diabetes, were found to be more susceptible to
respiratory failure and therefore, had poorer
prognoses. Providing respiratory support at early
stages improved the disease prognosis and facilitated
recovery (18). The ARDS in COVID-19 is due to the
occurrence of cytokine storms that results in
exaggerated immune response, immune regulatory
network imbalance, and, finally, multiple-organ
failure (122). In addition to the exaggerated
inflammatory response seen in patients with
COVID-19 pneumonia, the bile duct epithelial cell-
derived hepatocytes upregulate ACE2 expression in
liver tissue by compensatory proliferation that might
result in hepatic tissue injury (123).
CORONAVIRUSES IN ANIMALS AND
ZOONOTIC LINKS-A BRIEF
VIEWPOINT
Coronavirus can cause disease in several species
of domestic and wild animals, as well as humans
(23). The different animal species that are infected
with CoV include horses, camels, cattle, swine, dogs,
cats, rodents, birds, ferrets, minks, bats, rabbits,
snakes, and various other wild animals (20, 30, 79,
snakes, and various other wild animals (20, 30, 79,
93, 124, 125, 287). Coronavirus infection is linked to
different kinds of clinical manifestations, varying
from enteritis in cows and pigs, upper respiratory
disease in chickens, and fatal respiratory infections
in humans (30).
Among the CoV genera, Alphacoronavirus and
Betacoronavirus infect mammals, while
Gammacoronavirus and Deltacoronavirus mainly
infect birds, fishes, and, sometimes, mammals (27,
29, 106). Several novel coronaviruses that come
under the genus Deltacoronavirus have been
discovered in the past from birds, like Wigeon
coronavirus HKU20, Bulbul coronavirus HKU11,
Munia coronavirus HKU13, white-eye coronavirus
HKU16, night-heron coronavirus HKU19, and
common moorhen coronavirus HKU21, aswell as
from pigs (porcine coronavirus HKU15) (6, 29).
Transmissible gastroenteritis virus (TGEV), porcine
epidemic diarrhea virus (PEDV), and porcine
hemagglutinating encephalomyelitis virus (PHEV)
are some of the coronaviruses of swine. Among
them, TGEV and PEDV are responsible for causing
severe gastroenteritis in young piglets with
noteworthy morbidity and mortality. Infection with
PHEV also causes enteric infection but can cause
encephalitis due to its ability to infect the nervous
system (30).
Bovine coronaviruses (BoCoVs) are known to
infect several domestic and wild ruminants (126).
BoCoV inflicts neonatal calf diarrhea in adult cattle,
leading to bloody diarrhea (winter dysentery) and
respiratory disease complex (shipping fever) in cattle
of all age groups (126). BoCoV-like viruses have
been noted in humans, suggesting its zoonotic
potential as well (127). Feline enteric and feline
infectious peritonitis (FIP) viruses are the two major
feline CoVs (128), where feline CoVs can affect the
gastrointestinal tract, abdominal cavity (peritonitis),
respiratory tract, and central nervous system (128).
Canines are also affected by CoVs that fall under
different genera, namely, canine enteric coronavirus
in Alphacoronavirus and canine respiratory
coronavirus in Betacoronavirus, affecting the enteric
and respiratory tract, respectively (129, 130). IBV,
under Gammacoronavirus, causes diseases of
respiratory, urinary, and reproductive systems, with
substantial economic losses in chickens (131, 132).
In small laboratory animals, mouse hepatitis virus,
rat sialodacryoadenitis coronavirus, and guinea pig
and rabbit coronaviruses are the major CoVs
associated with disease manifestations like enteritis,
hepatitis, and respiratory infections (10, 133).
Swine acute diarrhea syndrome coronavirus
Swine acute diarrhea syndrome coronavirus
(SADS-CoV) was first identified in suckling piglets
having severe enteritis and belongs to the genus
Alphacoronavirus (106). The outbreak was
associated with considerable scale mortality of
piglets (24,693 deaths) across four farms in China
(134). The virus isolated from the piglets was almost
identical to and had 95% genomic similarity with
horseshoe bat (Rhinolophus species) coronavirus
HKU2, suggesting a bat origin of the pig virus (106,
134, 135). It is also imperative to note that the
SADS-CoV outbreak started in Guangdong province,
near the location of the SARS pandemic origin
(134). Before this outbreak, pigs were not known to
be infected with bat-origin coronaviruses. This
indicates that the bat-origin coronavirus jumped to
pig by breaking the species barrier. The next step of
this jump might not end well, since pigs are
considered the mixing vessel for influenza A viruses
due to their ability to be infected by both human and
avian influenza A viruses (136).
Similarly, they may act as the mixing vessel for
coronaviruses, since they are in frequent contact with
both humans and multiple wildlife species.
Additionally, pigs are also found to be susceptible to
infection with human SARS-CoV and MERS-CoV,
making this scenario a nightmare (109, 137). It is
only a matter of time before another zoonotic
coronavirus results in an epidemic by jumping the
so-called species barrier (287).
The host spectrum of coronavirus increased when
a novel coronavirus, namely, SW1, was recognized
in the liver tissue of a captive beluga whale
(Delphinapterus leucas) (138). In recent decades,
several novel coronaviruses were identified from
different animal species. Bats can harbor these
viruses without manifesting any clinical disease but
are persistently infected (30). They are the only
mammals with the capacity for self-powered flight,
which enables them to migrate long distances, unlike
land mammals. Bats are distributed worldwide and
also account for about a fifth of all mammalian
species (6). This makes them the ideal reservoir host
for many viral agents and also the source of novel
coronaviruses that have yet to be identified. It has
become a necessity to study the diversity of
coronavirus in the bat population to prevent future
outbreaks that could jeopardize livestock and public
health. The repeated outbreaks caused by bat-origin
coronaviruses calls for the development of efficient
molecular surveillance strategies for studying
Betacoronavirus among animals (12), especially in
the Rhinolophus bat family (86). Chinese bats have
high commercial value, since they are used in
high commercial value, since they are used in
traditional Chinese medicine (TCM). Therefore, the
handling of bats for trading purposes poses a
considerable risk of transmitting zoonotic CoV
epidemics (139)
Due to the possible role played by farm and wild
animals in SARS-CoV-2 infection, the WHO, in
their novel coronavirus (COVID-19) situation report,
recommended the avoidance of unprotected contact
with both farm and wild animals (25). The live-
animal markets, like the one in Guangdong, China,
provides a setting for animal coronaviruses to
amplify and to be transmitted to new hosts, like
humans (78). Such markets can be considered a
critical place for the origin of novel zoonotic
diseases and have enormous public health
significance in the event of an outbreak. Bats are the
reservoirs for several viruses; hence, the role of bats
in the present outbreak cannot be ruled out (140). In
a qualitative study conducted for evaluating the
zoonotic risk factors among rural communities of
southern China, the frequent human-animal
interactions along with the low levels of
environmental biosecurity were identified as
significant risks for the emergence of zoonotic
disease in local communities (141, 142).
The comprehensive sequence analysis of the
The comprehensive sequence analysis of the
SARS-CoV-2 RNA genome identified that the CoV
from Wuhan is a recombinant virus of the bat
coronavirus and another coronavirus of unknown
origin. The recombination was found to have
happened within the viral spike glycoprotein, which
recognizes the cell surface receptor. Further analysis
of the genome based on codon usage identified the
snake as the most probable animal reservoir of
SARS-CoV-2 (143). Contrary to these findings,
another genome analysis proposed that the genome
of SARS-CoV-2 is 96% identical to bat coronavirus,
reflecting its origin from bats (63). The involvement
of bat-derived materials in causing the current
outbreak cannot be ruled out. High risk is involved
in the production of bat-derived materials for TCM
practices involving the handling of wild bats. The
use of bats for TCM practices will remain a severe
risk for the occurrence of zoonotic coronavirus
epidemics in the future (139).
Furthermore, the pangolins are an endangered
species of animals that harbor a wide variety of
viruses, including coronaviruses (144). The
coronavirus isolated from Malayan pangolins (Manis
javanica) showed a very high amino acid identity
with COVID-19 at E (100%), M (98.2%), N
(96.7%), and S genes (90.4%). The RBD of S protein
(96.7%), and S genes (90.4%). The RBD of S protein
in CoV isolated from pangolin was almost identical
(one amino acid difference) to that of SARS-CoV-2.
A comparison of the genomes suggests
recombination between pangolin-CoV-like viruses
with the bat-CoV-RaTG13-like virus. All this
suggests the potential of pangolins to act as the
intermediate host of SARS-CoV-2 (145).
Human-wildlife interactions, which are
increasing in the context of climate change (142), are
further considered high risk and responsible for the
emergence of SARS-CoV. COVID-19 is also
suspected of having a similar mode of origin. Hence,
to prevent the occurrence of another zoonotic
spillover (1), exhaustive coordinated efforts are
needed to identify the high-risk pathogens harbored
by wild animal populations, conducting surveillance
among the people who are susceptible to zoonotic
spillover events (12), and to improve the biosecurity
measures associated with the wildlife trade (146).
The serological surveillance studies conducted in
people living in proximity to bat caves had earlier
identified the serological confirmation of SARS-
related CoVs in humans. People living at the
wildlife-human interface, mainly in rural China, are
regularly exposed to SARS-related CoVs (147).
These findings will not have any significance until a
These findings will not have any significance until a
significant outbreak occurs due to a virus-like
SARS-CoV-2.
There is a steady increase in the reports of
COVID-19 in companion and wild animals around
the world. Further studies are required to evaluate
the potential of animals (especially companion
animals) to serve as an efficient reservoir host that
can further alter the dynamics of human-to-human
transmission (330). To date, two pet dogs (Hong
Kong) and four pet cats (one each from Belgium and
Hong Kong, two from the United States) have tested
positive for SARS-CoV-2 (335). The World
Organization for Animal Health (OIE) has confirmed
the diagnosis of COVID-19 in both dogs and cats
due to human-to-animal transmission (331). The
similarity observed in the gene sequence of SARS-
CoV-2 from an infected pet owner and his dog
further confirms the occurrence of human-to-animal
transmission (333). Even though asymptomatic,
feline species should be considered a potential
transmission route from animals to humans (326).
However, currently, there are no reports of SARS-
CoV-2 transmission from felines to human beings.
Based on the current evidence, we can conclude that
cats are susceptible to SARS-CoV-2 and can get
infected by human beings. However, evidence of cat
infected by human beings. However, evidence of cat-
to-human transmission is lacking and requires
further studies (332). Rather than waiting for firmer
evidence on animal-to-human transmission,
necessary preventive measures are advised, as well
as following social distancing practices among
companion animals of different households (331).
One of the leading veterinary diagnostic companies,
IDEXX, has conducted large-scale testing for
COVID-19 in specimens collected from dogs and
cats. However, none of the tests turned out to be
positive (334).
In a study conducted to investigate the potential
of different animal species to act as the intermediate
host of SARS-CoV-2, it was found that both ferrets
and cats can be infected via experimental inoculation
of the virus. In addition, infected cats efficiently
transmitted the disease to naive cats (329). SARS-
CoV-2 infection and subsequent transmission in
ferrets were found to recapitulate the clinical aspects
of COVID-19 in humans. The infected ferrets also
shed virus via multiple routes, such as saliva, nasal
washes, feces, and urine, postinfection, making them
an ideal animal model for studying disease
transmission (337). Experimental inoculation was
also done in other animal species and found that the
dogs have low susceptibility, while the chickens,
dogs have low susceptibility, while the chickens,
ducks, and pigs are not at all susceptible to SARS-
CoV-2 (329).
Similarly, the National Veterinary Services
Laboratories of the USDA have reported COVID-19
in tigers and lions that exhibited respiratory signs
like dry cough and wheezing. The zoo animals are
suspected to have been infected by an asymptomatic
zookeeper (335). The total number of COVID-19-
positive cases in human beings is increasing at a high
rate, thereby creating ideal conditions for viral
spillover to other species, such as pigs. The evidence
obtained from SARS-CoV suggests that pigs can get
infected with SARS-CoV-2 (336). However,
experimental inoculation with SARS-CoV-2 failed to
infect pigs (329).
Further studies are required to identify the
possible animal reservoirs of SARS-CoV-2 and the
seasonal variation in the circulation of these viruses
in the animal population. Research collaboration
between human and animal health sectors is
becoming a necessity to evaluate and identify the
possible risk factors of transmission between animals
and humans. Such cooperation will help to devise
efficient strategies for the management of emerging
zoonotic diseases (12).
DIAGNOSIS OF SARS-CoV-2 (COVID-
19)
RNA tests can confirm the diagnosis of SARS-
CoV-2 (COVID-19) cases with real-time RT-PCR or
next-generation sequencing (148, 149, 245, 246). At
present, nucleic acid detection techniques, like RT-
PCR, are considered an effective method for
confirming the diagnosis in clinical cases of COVID-
19 (148). Several companies across the world are
currently focusing on developing and marketing
SARS-CoV-2-specific nucleic acid detection kits.
Multiple laboratories are also developing their own
in-house RT-PCR. One of them is the SARS-CoV-2
nucleic acid detection kit produced by Shuoshi
Biotechnology (double fluorescence PCR method)
(150). Up to 30 March 2020, the U.S. Food and Drug
Administration (FDA) had granted 22 in vitro
diagnostics Emergency Use Authorizations (EUAs),
including for the RT-PCR diagnostic panel for the
universal detection of SARS-like betacoronaviruses
and specific detection of SARS-CoV-2, developed
by the U.S. CDC (Table 1) (258, 259).
Recently, 95 full-length genomic sequences of
SARAS-CoV-2 strains available in the National
Center for Biotechnology Information and GISAID
databases were subjected to multiple-sequence
alignment and phylogenetic analyses for studying
variations in the viral genome (260). All the viral
strains revealed high homology of 99.99% (99.91%
to 100%) at the nucleotide level and 99.99%
(99.79% to 100%) at the amino acid level. Overall
variation was found to be low in ORF regions, with
13 variation sites recognized in 1a, 1b, S, 3a, M, 8,
and N regions. Mutation rates of 30.53% (29/95)
and 29.47% (28/95) were observed at nt 28144 (ORF8)
and nt 8782 (ORFla) positions, respectively. Owing
to such selective mutations, a few specific regions of
SARS-CoV-2 should not be considered for designing
primers and probes. The SARS-CoV-2 reference
sequence could pave the way to study molecular
biology and pathobiology, along with developing
diagnostics and appropriate prevention and control
strategies for countering SARS-CoV-2 (260).
Nucleic acids of SARS-CoV-2 can be detected
from samples (64) such as bronchoalveolar lavage
fluid, sputum, nasal swabs, fiber bronchoscope brush
biopsy specimen, pharyngeal swabs, feces, blood,
and urine, with different levels of diagnostic
performance (Table 2) (80, 245, 246). The viral loads
performance (Table 2) (80, 245, 246). The viral loads
of SARS-CoV-2 were measured using N-gene-
specific quantitative RT-PCR in throat swab and
sputum samples collected from COVID-19-infected
individuals. The results indicated that the viral load
peaked at around 5 to 6 days following the onset of
symptoms, and it ranged from 104 to 10' copies/ml
during this time (151). In another study, the viral
load was found to be higher in the nasal swabs than
the throat swabs obtained from COVID-19
symptomatic patients (82). Although initially it was
thought that viral load would be associated with poor
outcomes, some case reports have shown
asymptomatic individuals with high viral loads
(247). Recently, the viral load in nasal and throat
swabs of 17 symptomatic patients was determined,
and higher viral loads were recorded soon after the
onset of symptoms, particularly in the nose
compared to the throat. The pattern of viral nucleic
acid shedding of SARS-CoV-2-infected patients was
similar to that of influenza patients but seemed to be
different from that of SARS-CoV patients. The viral
load detected in asymptomatic patients resembled
that of symptomatic patients as studied in China,
which reflects the transmission perspective of
asymptomatic or symptomatic patients having
minimum signs and symptoms (82). Another study,
minimum signs and symptoms (82). Another study,
conducted in South Korea, related to SARS-CoV-2
viral load, opined that SARS-CoV-2 kinetics were
significantly different from those of earlier reported
CoV infections, including SARS-CoV (253). SARS-
CoV-2 transmission can occur early in the viral
infection phase; thus, diagnosing cases and isolation
attempts for this virus warrant different strategies
than those needed to counter SARS-CoV. Studies are
required to establish any correlation between SARS-
CoV-2 viral load and cultivable virus. Recognizing
patients with fewer or no symptoms, along with
having modest detectable viral RNA in the
oropharynx for 5 days, indicates the requirement of
data for assessing SARS-CoV-2 transmission
dynamics and updating the screening procedures in
the clinics (82)
The results of the studies related to SARS-CoV-2
viral loads reflect active replication of this virus in
the upper respiratory tract and prolonged viral
shedding after symptoms disappear, including via
stool. Thus, the current case definition needs to be
updated along with a reassessment of the strategies
to be adopted for restraining the SARS-CoV-2
outbreak spread (248). In some cases, the viral load
studies of SARS-CoV-2 have also been useful to
recommend precautionary measures when handling
specific samples, e.g., feces. In a recent survey from
17 confirmed cases of SARS-CoV-2 infection with
available data (representing days 0 to 13 after onset),
stool samples from nine cases (53%; days 0 to 11
after onset) were positive on RT-PCR analysis.
Although the viral loads were lower than those of
respiratory samples (range, 550 copies per ml to
1.21 x 10 copies per ml), this has essential biosafety
implications (151).
The samples from 18 SARS-CoV-2-positive
patients in Singapore who had traveled from Wuhan
to Singapore showed the presence of viral RNA in
stool and whole blood but not in urine by real-time
RT-PCR (288). Further, novel SARS-CoV-2
infections have been detected in a variety of clinical
specimens, like bronchoalveolar lavage fluid,
specimens, like bronchoalveolar lavage fluid,
sputum, nasal swabs, fibrobronchoscope brush
biopsy specimens, pharyngeal swabs, feces, and
blood (246).
The presence of SARS-CoV-2 in fecal samples
has posed grave public health concerns. In addition
to the direct transmission mainly occurring via
droplets of sneezing and coughing, other routes, such
as fecal excretion and environmental and fomite
contamination, are contributing to SARS-CoV-2
transmission and spread (249–252). Fecal excretion
has also been documented for SARS-CoV and
MERS-CoV, along with the potential to stay viable
in situations aiding fecal-oral transmission. Thus,
SARS-CoV-2 has every possibility to be transmitted
through this mode. Fecal-oral transmission of SARS-
CoV-2, particularly in regions having low standards
of hygiene and poor sanitation, may have grave
consequences with regard to the high spread of this
virus. Ethanol and disinfectants containing chlorine
or bleach are effective against coronaviruses
(249–252). Appropriate precautions need to be
followed strictly while handling the stools of patients
infected with SARS-CoV-2. Biowaste materials and
sewage from hospitals must be adequately
disinfected, treated, and disposed of properly. The
significance of frequent and good hand hygiene and
significance of frequent and good hand hygiene and
sanitation practices needs to be given due emphasis
(249–252). Future explorative research needs to be
conducted with regard to the fecal-oral transmission
of SARS-CoV-2, along with focusing on
environmental investigations to find out if this virus
could stay viable in situations and atmospheres
facilitating such potent routes of transmission. The
correlation of fecal concentrations of viral RNA with
disease severity needs to be determined, along with
assessing the gastrointestinal symptoms and the
possibility of fecal SARS-CoV-2 RNA detection
during the COVID-19 incubation period or
convalescence phases of the disease
(249–252).
The lower respiratory tract sampling techniques,
like bronchoalveolar lavage fluid aspirate, are
considered the ideal clinical materials, rather than
the throat swab, due to their higher positive rate on
the nucleic acid test (148). The diagnosis of COVID-
19 can be made by using upper-respiratory-tract
specimens collected using nasopharyngeal and
oropharyngeal swabs. However, these techniques are
associated with unnecessary risks to health care
workers due to close contact with patients (152).
Similarly, a single patient with a high viral load was
reported to contaminate an entire endoscopy room by
shedding the virus, which may remain viable for at
shedding the virus, which may remain viable for at
least 3 days and is considered a great risk for
uninfected patients and health care workers (289).
Recently, it was found that the anal swabs gave more
positive results than oral swabs in the later stages of
infection (153). Hence, clinicians have to be cautious
while discharging any COVID-19-infected patient
based on negative oral swab test results due to the
possibility of fecal-oral transmission. Even though
the viral loads in stool samples were found to be less
than those of respiratory samples, strict
precautionary measures have to be followed while
handling stool samples of COVID-19 suspected or
infected patients (151). Children infected with
SARS-CoV-2 experience only a mild form of illness
and recover immediately after treatment. It was
recently found that stool samples of SARS-CoV-2-
infected children that gave negative throat swab
results were positive within ten days of negative
results. This could result in the fecal-oral
transmission of SARS-CoV-2 infections, especially
in children (290). Hence, to prevent the fecal-oral
transmission of SARS-CoV-2, infected COVID-19
patients should only be considered negative when
they test negative for SARS-CoV-2 in the stool
sample.
A suspected case of COVID-19 infection is said
to be confirmed if the respiratory tract aspirate or
blood samples test positive for SARS-CoV-2 nucleic
acid using RT-PCR or by the identification of SARS-
CoV-2 genetic sequence in respiratory tract aspirate
or blood samples (80). The patient will be confirmed
as cured when two subsequent oral swab results are
negative (153). Recently, the live virus was detected
in the self-collected saliva of patients infected with
COVID-19. These findings were confirmative of
using saliva as a noninvasive specimen for the
diagnosis of COVID-19 infection in suspected
individuals (152). It has also been observed that the
initial screening of COVID-19 patients infected with
RT-PCR may give negative results even if they have
chest CT findings that are suggestive of infection.
Hence, for the accurate diagnosis of COVID-19, a
combination of repeated swab tests using RT-PCR
and CT scanning is required to prevent the
possibility of false-negative results during disease
screening (154). RT-PCR is the most widely used test
for diagnosing COVID-19. However, it has some
significant limitations from the clinical perspective,
since it will not give any clarity regarding disease
progression. Droplet digital PCR (ddPCR) can be
used for the quantification of viral load in the
samples obtained from lower respiratory tracts.
samples obtained from lower respiratory tracts.
Hence, based on the viral load, we can quickly
evaluate the progression of infection (291). In
addition to all of the above findings, sequencing and
phylogenetics are critical in the correct identification
and confirmation of the causative viral agent and
useful to establish relationships with previous
isolates and sequences, as well as to know, especially
during an epidemic, the nucleotide and amino acid
mutations and the molecular divergence. The rapid
development and implementation of diagnostic tests
against emerging novel diseases like COVID-19
pose significant challenges due to the lack of
resources and logistical limitations associated with
an outbreak (155).
SARS-CoV-2 infection can also be confirmed by
isolation and culturing. The human airway epithelial
cell culture was found to be useful in isolating
SARS-CoV-2 (3). The efficient control of an
outbreak depends on the rapid diagnosis of the
disease. Recently, in response to the COVID-19
outbreak, 1-step quantitative real-time reverse
transcription-PCR assays were developed that detect
the ORF1b and N regions of the SARS-CoV-2
genome (156). That assay was found to achieve the
rapid detection of SARS-CoV-2. Nucleic acid-based
assays offer high accuracy in the diagnosis of SARS
assays offer high accuracy in the diagnosis of SARS-
CoV-2, but the current rate of spread limits its use
due to the lack of diagnostic assay kits. This will
further result in the extensive transmission of
COVID-19, since only a portion of suspected cases
can be diagnosed. In such situations, conventional
serological assays, like enzyme-linked
immunosorbent assay (ELISA), that are specific to
COVID-19 IgM and IgG antibodies can be used as a
high-throughput alternative (149). At present, there
is no diagnostic kit available for detecting the SARS-
CoV-2 antibody (150). The specific antibody profiles
of COVID-19 patients were analyzed, and it was
found that the IgM level lasted more than 1 month,
indicating a prolonged stage of virus replication in
SARS-CoV-2-infected patients. The IgG levels were
found to increase only in the later stages of the
disease. These findings indicate that the specific
antibody profiles of SARS-CoV-2 and SARS-CoV
were similar (325). These findings can be utilized for
the development of specific diagnostic tests against
COVID-19 and can be used for rapid screening.
Even though diagnostic test kits are already available
that can detect the genetic sequences of SARS-CoV-
2 (95), their availability is a concern, as the number
of COVID-19 cases is skyrocketing (155, 157). A
major problem associated with this diagnostic kit is
major problem associated with this diagnostic kit is
that it works only when the test subject has an active
infection, limiting its use to the earlier stages of
infection. Several laboratories around the world are
currently developing antibody-based diagnostic tests
against SARS-CoV-2 (157).
Chest CT is an ideal diagnostic tool for
identifying viral pneumonia. The sensitivity of chest
CT is far superior to that of X-ray screening. The
chest CT findings associated with COVID-19-
infected patients include characteristic patchy
infiltration that later progresses to ground-glass
opacities (158). Early manifestations of COVID-19
pneumonia might not be evident in X-ray chest
radiography. In such situations, a chest CT
examination can be performed, as it is considered
highly specific for COVID-19 pneumonia (118).
Those patients having COVID-19 pneumonia will
exhibit the typical ground-glass opacity in their chest
CT images (154). The patients infected with
COVID-19 had elevated plasma angiotensin 2 levels.
The level of angiotensin 2 was found to be linearly
associated with viral load and lung injury, indicating
its potential as a diagnostic biomarker (121). The
chest CT imaging abnormalities associated with
COVID-19 pneumonia have also been observed even
in asymptomatic patients. These abnormalities
in asymptomatic patients. These abnormalities
progress from the initial focal unilateral to diffuse
bilateral ground-glass opacities and will further
progress to or coexist with lung consolidation
changes within 1 to 3 weeks (159). The role played
by radiologists in the current scenario is very
important. Radiologists can help in the early
diagnosis of lung abnormalities associated with
COVID-19 pneumonia. They can also help in the
evaluation of disease severity, identifying its
progression to acute respiratory distress syndrome
and the presence of secondary bacterial infections
(160). Even though chest CT is considered an
essential diagnostic tool for COVID-19, the
extensive use of CT for screening purposes in the
suspected individuals might be associated with a
disproportionate risk-benefit ratio due to increased
radiation exposure as well as increased risk of cross-
infection. Hence, the use of CT for early diagnosis of
SARS-CoV-2 infection in high-risk groups should be
done with great caution (292).
More recently, other advanced diagnostics have
been designed and developed for the detection of
SARS-CoV-2 (345, 347, 350–352). A reverse
transcriptional loop-mediated isothermal
amplification (RT-LAMP), namely, iLACO, has been
developed for rapid and colorimetric detection of this
developed for rapid and colorimetric detection of this
virus (354). RT-LAMP serves as a simple, rapid, and
sensitive diagnostic method that does not require
sophisticated equipment or skilled personnel (349).
An interactive web-based dashboard for tracking
SARS-CoV-2 in a real-time mode has been designed
(238). A smartphone-integrated home-based point-
of-care testing (POCT) tool, a paper-based POCT
combined with LAMP, is a useful point-of-care
diagnostic (353). An Abbott ID Now COVID-19
molecular POCT-based test, using isothermal nucleic
acid amplification technology, has been designed as
a point-of-care test for very rapid detection of
SARS-CoV-2 in just 5 min (344). A CRISPR-based
SHERLOCK (specific high-sensitivity enzymatic
reporter unlocking) diagnostic for rapid detection of
SARS-CoV-2 without the requirement of specialized
instrumentation has been reported to be very useful
in the clinical diagnosis of COVID-19 (360). A
CRISPR-Cas 12-based lateral flow assay also has
been developed for rapid detection of SARS-CoV-2
(346). Artificial intelligence, by means of a three-
dimensional deep-learning model, has been
developed for sensitive and specific diagnosis of
COVID-19 via CT images (332).
Tracking and mapping of the rising incidence
rates, disease outbreaks, community spread,
rates, disease outbreaks, community spread,
clustered transmission events, hot spots, and
superspreader potential of SARS-CoV-2/COVID
warrant full exploitation of real-time disease
mapping by employing geographical information
systems (GIS), such as the GIS software Kosmo 3.1,
web-based real-time tools and dashboards, apps, and
advances in information technology (356–359).
Researchers have also developed a few prediction
tools/models, such as the prediction model risk of
bias assessment tool (PROBAST) and critical
appraisal and data extraction for systematic reviews
of prediction modeling studies (CHARMS), which
could aid in assessing the possibility of getting
infection and estimating the prognosis in patients;
however, such models may suffer from bias issues
and, hence, cannot be considered completely
trustworthy, which necessitates the development of
new and reliable predictors (360).
VACCINES, THERAPEUTICS, AND
DRUGS
Recently emerged viruses, such as Zika, Ebola,
and Nipah viruses, and their grave threats to humans
have begun a race in exploring the designing and
developing of advanced vaccines, prophylactics,
therapeutics, and drug regimens to counter emerging
therapeutics, and drug regimens to counter emerging
viruses (161–163, 280). Several attempts are being
made to design and develop vaccines for CoV
infection, mostly by targeting the spike glycoprotein.
Nevertheless, owing to extensive diversity in
antigenic variants, cross-protection rendered by the
vaccines is significantly limited, even within the
strains of a phylogenetic subcluster (104). Due to the
lack of effective antiviral therapy and vaccines in the
present scenario, we need to depend solely on
implementing effective infection control measures to
lessen the risk of possible nosocomial transmission
(68). Recently, the receptor for SARS-CoV-2 was
established as the human angiotensin-converting
enzyme 2 (hACE2), and the virus was found to enter
the host cell mainly through endocytosis. It was also
found that the major components that have a critical
role in viral entry include PIKfyve, TPC2, and
cathepsin L. These findings are critical, since the
components described above might act as candidates
for vaccines or therapeutic drugs against SARS-
CoV-2 (293).
The majority of the treatment options and
strategies that are being evaluated for SARS-CoV-2
(COVID-19) have been taken from our previous
experiences in treating SARS-CoV, MERS-CoV, and
other emerging viral diseases. Several therapeutic
other emerging viral diseases. Several therapeutic
and preventive strategies, including vaccines,
immunotherapeutics, and antiviral drugs, have been
exploited against the previous CoV outbreaks
(SARS-CoV and MERS-CoV) (8, 104, 164–167).
These valuable options have already been evaluated
for their potency, efficacy, and safety, along with
several other types of current research that will fuel
our search for ideal therapeutic agents against
COVID-19 (7, 9, 19, 21, 36). The primary cause of
the unavailability of approved and commercial
vaccines, drugs, and therapeutics to counter the
earlier SARS-CoV and MERS-CoV seems to
owe to the lesser attention of the biomedicine and
pharmaceutical companies, as these two CoVs did
not cause much havoc, global threat, and panic like
those posed by the SARS-CoV-2 pandemic (19).
Moreover, for such outbreak situations, the
requirement for vaccines and therapeutics/drugs
exists only for a limited period, until the outbreak is
controlled. The proportion of the human population
infected with SARS-CoV and MERS-CoV was also
much lower across the globe, failing to attract drug
and vaccine manufacturers and producers. Therefore
by the time an effective drug or vaccine is designed
against such disease outbreaks, the virus would have
been controlled by adopting appropriate and strict
been controlled by adopting appropriate and strict
prevention and control measures, and patients for
clinical trials will not be available. The newly
developed drugs cannot be marketed due to the lack
of end users.
Vaccines
The S protein plays a significant role in the
induction of protective immunity against SARS-CoV
by mediating T-cell responses and neutralizing
antibody production (168). In the past few decades,
we have seen several attempts to develop a vaccine
against human coronaviruses by using S protein as
the target (168, 169). However, the developed
vaccines have minimal application, even among
closely related strains of the virus, due to a lack of
cross-protection. That is mainly because of the
extensive diversity existing among the different
antigenic variants of the virus (104). The
contributions of the structural proteins, like spike
(S), matrix (M), small envelope (E), and
nucleocapsid (N) proteins, of SARS-CoV to induce
protective immunity has been evaluated by
expressing them in a recombinant parainfluenza
virus type 3 vector (BHPIV3). Of note, the result
was conclusive that the expression of M, E, or N
proteins without the presence of S protein would not
proteins without the presence of S protein would not
confer any noticeable protection, with the absence of
detectable serum SARS-CoV-neutralizing antibodies
(170). Antigenic determinant sites present over S and
N structural proteins of SARS-CoV-2 can be
explored as suitable vaccine candidates (294). In the
Asian population, S, E, M, and N proteins of SARS-
CoV-2 are being targeted for developing subunit
vaccines against COVID-19 (295).
The identification of the immunodominant region
among the subunits and domains of S protein is
critical for developing an effective vaccine against
the coronavirus. The C-terminal domain of the S1
subunit is considered the immunodominant region of
the porcine deltacoronavirus S protein (171).
Similarly, further investigations are needed to
determine the immunodominant regions of SARS-
CoV-2 for facilitating vaccine development.
However, our previous attempts to develop a
universal vaccine that is effective for both SARS-
CoV and MERS-CoV based on T-cell epitope
similarity pointed out the possibility of cross-
reactivity among coronaviruses (172). That can be
made possible by selected potential vaccine targets
that are common to both viruses. SARS-CoV-2 has
been reported to be closely related to SARS-CoV
(173, 174). Hence, knowledge and understanding of
(173, 174). Hence, knowledge and understanding of
S protein-based vaccine development in SARS-CoV
will help to identify potential S protein vaccine
candidates in SARS-CoV-2. Therefore, vaccine
strategies based on the whole S protein, S protein
subunits, or specific potential epitopes of S protein
appear to be the most promising vaccine candidates
against coronaviruses. The RBD of the S1 subunit of
S protein has a superior capacity to induce
neutralizing antibodies. This property of the RBD
can be utilized for designing potential SARS-CoV
vaccines either by using RBD-containing
recombinant proteins or recombinant vectors that
encode RBD (175). Hence, the superior genetic
similarity existing between SARS-CoV-2 and SARS-
CoV can be utilized to repurpose vaccines that have
proven in vitro efficacy against SARS-CoV to be
utilized for SARS-CoV-2. The possibility of cross-
protection in COVID-19 was evaluated by
comparing the S protein sequences of SARS-CoV-2
with that of SARS-CoV. The comparative analysis
confirmed that the variable residues were found
concentrated on the S1 subunit of S protein, an
important vaccine target of the virus (150). Hence,
the possibility of SARS-CoV-specific neutralizing
antibodies providing cross-protection to COVID-19
might be lower. Further genetic analysis is required
might be lower. Further genetic analysis is required
between SARS-CoV-2 and different strains of
SARS-CoV and SARS-like (SL) COVs to evaluate
the possibility of repurposed vaccines against
COVID-19. This strategy will be helpful in the
scenario of an outbreak, since much time can be
saved, because preliminary evaluation, including in
vitro studies, already would be completed for such
vaccine candidates.
Multiepitope subunit vaccines can be considered
a promising preventive strategy against the ongoing
COVID-19 pandemic. In silico and advanced
immunoinformatic tools can be used to develop
multiepitope subunit vaccines. The vaccines that are
engineered by this technique can be further evaluated
using docking studies and, if found effective, then
can be further evaluated in animal models (365).
Identifying epitopes that have the potential to
become a vaccine candidate is critical to developing
an effective vaccine against COVID-19. The
immunoinformatics approach has been used for
recognizing essential epitopes of cytotoxic T
lymphocytes and B cells from the surface
glycoprotein of SARS-CoV-2. Recently, a few
epitopes have been recognized from the SARS-CoV-
2 surface glycoprotein. The selected epitopes
explored targeting molecular dynamic simulations,
explored targeting molecular dynamic simulations,
evaluating their interaction with corresponding major
histocompatibility complex class I molecules. They
potentially induce immune responses (176). The
recombinant vaccine can be designed by using rabies
virus (RV) as a viral vector. RV can be made to
express MERS-CoV S1 protein on its surface so that
an immune response is induced against MERS-CoV.
The RV vector-based vaccines against MERS-CoV
can induce faster antibody response as well as higher
degrees of cellular immunity than the Gram-positive
enhancer matrix (GEM) particle vector-based
vaccine. However, the latter can induce a very high
antibody response at lower doses (167). Hence, the
degree of humoral and cellular immune responses
produced by such vaccines depends upon the vector
used.
Dual vaccines have been getting more popular
recently. Among them, the rabies virus-based
vectored vaccine platform is used to develop
vaccines against emerging infectious diseases. The
dual vaccine developed from inactivated rabies virus
particles that express the MERS-CoV S1 domain of
S protein was found to induce immune responses for
both MERS-CoV and rabies virus. The vaccinated
mice were found to be completely protected from
challenge with MERS-CoV (169). The intranasal
administration of the recombinant adenovirus-based
vaccine in BALB/c mice was found to induce long-
lasting neutralizing immunity against MERS spike
pseudotyped virus, characterized by the induction of
systemic IgG, secretory IgA, and lung-resident
memory T-cell responses (177). Immunoinformatics
methods have been employed for the genome-wide
screening of potential vaccine targets among the
different immunogens of MERS-CoV (178). The N
protein and the potential B-cell epitopes of MERS-
CoV E protein have been suggested as
immunoprotective targets inducing both T-cell and
neutralizing antibody responses (178, 179).
The collaborative effort of the researchers of
Rocky Mountain Laboratories and Oxford University
is designing a chimpanzee adenovirus-vectored
vaccine to counter COVID-19 (180). The Coalition
for Epidemic Preparedness Innovations (CEPI) has
initiated three programs to design SARS-CoV-2
vaccines (181). CEPI has a collaborative project with
Inovio for designing a MERS-CoV DNA vaccine
that could potentiate effective immunity. CEPI and
the University of Queensland are designing a
molecular clamp vaccine platform for MERS-CoV
and other pathogens, which could assist in the easier
identification of antigens by the immune system
(181), CEPI has also funded Moderna to develop a
(181). CEPI has also funded Moderna to develop a
vaccine for COVID-19 in partnership with the
Vaccine Research Center (VRC) of the National
Institute of Allergy and Infectious Diseases (NIAID),
part of the National Institutes of Health (NIH) (182).
By employing mRNA vaccine platform technology, a
vaccine candidate expressing SARS-CoV-2 spike
protein is likely to go through clinical testing in the
coming months (180). On 16 March 2020, Jennifer
Haller became the first person outside China to
receive an experimental vaccine, developed by
Moderna, against this pandemic virus. Moderna,
along with China's CanSino Biologics, became the
first research group to launch small clinical trials of
vaccines against COVID-19. Their study is
evaluating the vaccine’s safety and ability to trigger
immune responses (296).
Scientists from all over the world are trying hard
to develop working vaccines with robust protective
immunity against COVID-19. Vaccine candidates,
like mRNA-1273 SARS-CoV-2 vaccine, INO-4800
DNA coronavirus vaccine, and adenovirus type 5
vector vaccine candidate (Ad5-nCoV), are a few
examples under phase I clinical trials, while self-
amplifying RNA vaccine, oral recombinant COVID-
19 vaccine, BNT162, plant-based COVID-19
vaccine, and li-Key peptide COVID-19 vaccine are
vaccine, and li-Key peptide COVID-19 vaccine are
under preclinical trials (297). Similarly, the WHO,
on its official website, has mentioned a detailed list
of COVID-19 vaccine agents that are under
consideration. Different phases of trials are ongoing
for live attenuated virus vaccines, formaldehyde
alum inactivated vaccine, adenovirus type 5 vector
vaccine, LNP-encapsulated mRNA vaccine, DNA
plasmid vaccine, and S protein, S-trimer, and Ii-Key
peptide as a subunit protein vaccine, among others
(298). The process of vaccine development usually
takes approximately ten years, in the case of
inactivated or live attenuated vaccines, since it
involves the generation of long-term efficacy data.
However, this was brought down to 5 years during
the Ebola emergency for viral vector vaccines. In the
urgency associated with the COVID-19 outbreaks,
we expect a vaccine by the end of this year (343).
The development of an effective vaccine against
COVID-19 with high speed and precision is the
combined result of advancements in computational
biology, gene synthesis, protein engineering, and the
invention of advanced manufacturing platforms (342).
The recurring nature of the coronavirus outbreaks
calls for the development of a pan-coronavirus
vaccine that can produce cross-reactive antibodies.
vaccine that can produce cross-reactive antibodies.
However, the success of such a vaccine relies greatly
on its ability to provide protection not only against
present versions of the virus but also the ones that
are likely to emerge in the future. This can be
achieved by identifying antibodies that can recognize
relatively conserved epitopes that are maintained as
such even after the occurrence of considerable
variations (362). Even though several vaccine
clinical trials are being conducted around the world,
pregnant women have been completely excluded
from these studies. Pregnant women are highly
vulnerable to emerging diseases such as COVID-19
due to alterations in the immune system and other
physiological systems that are associated with
pregnancy. Therefore, in the event of successful
vaccine development, pregnant women will not get
access to the vaccines (361). Hence, it is
recommended that pregnant women be included in
the ongoing vaccine trials, since successful
vaccination in pregnancy will protect the mother,
fetus, and newborn.
The heterologous immune effects induced by
Bacillus Calmette Guérin (BCG) vaccination is a
promising strategy for controlling the COVID-19
pandemic and requires further investigations. BCG is
a widely used vaccine against tuberculosis in high
risk regions. It is derived from a live attenuated
strain of Mycobacterium bovis. At present, three new
clinical trials have been registered to evaluate the
protective role of BCG vaccination against SARS-
COV-2 (363). Recently, a cohort study was conducted
to evaluate the impact of childhood BCG vaccination
in COVID-19 PCR positivity rates. However,
childhood BCG vaccination was found to be
associated with a rate of COVID-19-positive test
results similar to that of the nonvaccinated group
(364). Further studies are required to analyze
whether BCG vaccination in childhood can induce
protective effects against COVID-19 in adulthood.
Population genetic studies conducted on 103
genomes identified that the SARS-CoV-2 virus has
evolved into two major types, L and S. Among the
two types, L type is expected to be the most
prevalent (-70%), followed by the S type (-30%)
(366). This finding has a significant impact on our
race to develop an ideal vaccine, since the vaccine
candidate has to target both strains to be considered
effective. At present, the genetic differences between
the L and S types are very small and may not affect
the immune response. However, we can expect
further genetic variations in the coming days that
could lead to the emergence of new strains (367).
Therapeutics and Drugs
There is no currently licensed specific antiviral
treatment for MERS- and SARS-CoV infections, and
the main focus in clinical settings remains on
lessening clinical signs and providing supportive
care (183–186). Effective drugs to manage COVID-
19 patients include remdesivir, lopinavir/ritonavir
alone or in a blend with interferon beta, convalescent
plasma, and monoclonal antibodies (MAbs);
however, efficacy and safety issues of these drugs
require additional clinical trials (187, 281). A
controlled trial of ritonavir-boosted lopinavir and
interferon alpha 2b treatment was performed on
COVID-19 hospitalized patients
(ChiCTR2000029308) (188). In addition, the use of
hydroxychloroquine and tocilizumab for their
potential role in modulating inflammatory responses
in the lungs and antiviral effect has been proposed
and discussed in many research articles. Still, no
fool-proof clinical trials have been published (194,
196, 197, 261–272). Recently, a clinical trial
conducted on adult patients suffering from severe
COVID-19 revealed no benefit of lopinavir-ritonavir
treatment over standard care (273).
The efforts to control SARS-CoV-2 infection
utilize defined strategies as followed against MERS
and SARS, along with adopting and strengthening a
and SARS, along with adopting and strengthening a
few precautionary measures owing to the unknown
nature of this novel virus (36, 189). Presently, the
main course of treatment for severely affected
SARS-CoV-2 patients admitted to hospitals includes
mechanical ventilation, intensive care unit (ICU)
admittance, and symptomatic and supportive
therapies. Additionally, RNA synthesis inhibitors
(lamivudine and tenofovir disoproxil fumarate),
remdesivir, neuraminidase inhibitors, peptide (EK1),
anti-inflammatory drugs, abidol, and Chinese
traditional medicine (Lianhuaqingwen and
ShuFengJieDu capsules) could aid in COVID-19
treatment. However, further clinical trials are being
carried out concerning their safety and efficacy (7).
It might require months to a year(s) to design and
develop effective drugs, therapeutics, and vaccines
against COVID-19, with adequate evaluation and
approval from regulatory bodies and moving to the
bulk production of many millions of doses at
commercial levels to meet the timely demand of
mass populations across the globe (9). Continuous
efforts are also warranted to identify and assess
viable drugs and immunotherapeutic regimens that
revealed proven potency in combating other viral
agents similar to SARS-CoV-2.
COVID-19 patients showing severe signs are
COVID-19 patients showing severe signs are
treated symptomatically along with oxygen therapy.
In such cases where the patients progress toward
respiratory failure and become refractory to oxygen
therapy, mechanical ventilation is necessitated. The
COVID-19-induced septic shock can be managed by
providing adequate hemodynamic support (299).
Several classes of drugs are currently being
evaluated for their potential therapeutic action
against SARS-CoV-2. Therapeutic agents that have
anti-SARS-CoV-2 activity can be broadly classified
into three categories: drugs that block virus entry
into the host cell, drugs that block viral replication as
well as its survival within the host cell, and drugs
that attenuate the exaggerated host immune response
(300). An inflammatory cytokine storm is commonly
seen in critically ill COVID-19 patients. Hence, they
may benefit from the use of timely anti-inflammation
treatment. Anti-inflammatory therapy using drugs
like glucocorticoids, cytokine inhibitors, JAK
inhibitors, and chloroquine/hydroxychloroquine
should be done only after analyzing the risk/benefit
ratio in COVID-19 patients (301). There have not
been any studies concerning the application of
nonsteroidal anti-inflammatory drugs (NSAID) to
COVID-19-infected patients. However, reasonable
pieces of evidence are available that link NSAID
pieces of evidence are available that link NSAID
uses with the occurrence of respiratory and
cardiovascular adverse effects. Hence, as a
cautionary approach, it is better to recommend the
use of NSAIDs as the first-line option for managing
COVID-19 symptoms (302). The use of
corticosteroids in COVID-19 patients is still a matter
of controversy and requires further systematic
clinical studies. The guidelines that were put forward
to manage critically ill adults suggest the use of
systemic corticosteroids in mechanically ventilated
adults with ARDS (303). The generalized use of
corticosteroids is not indicated in COVID-19, since
there are some concerns associated with the use of
corticosteroids in viral pneumonia. Stem cell therapy
using mesenchymal stem cells (MSCs) is another
hopeful strategy that can be used in clinical cases of
COVID-19 owing to its potential
immunomodulatory capacity. It may have a
beneficial role in attenuating the cytokine storm that
is observed in severe cases of SARS-CoV-2
infection, thereby reducing mortality. Among the
different types of MSCs, expanded umbilical cord
MSCs can be considered a potential therapeutic
agent that requires further validation for managing
critically ill COVID-19 patients (304).
Repurposed broad-spectrum antiviral drugs
Repurposea broad-spectrum antiviral drugs
having proven uses against other viral pathogens can
be employed for SARS-CoV-2-infected patients.
These possess benefits of easy accessibility and
recognized pharmacokinetic and pharmacodynamics
activities, stability, doses, and side effects (9).
Repurposed drugs have been studied for treating
CoV infections, like lopinavir/ritonavir, and
interferon-1B revealed in vitro anti-MERS-CoV
action. The in vivo experiment carried out in the
nonhuman primate model of common marmosets
treated with lopinavir/ritonavir and interferon beta
showed superior protective results in treated animals
than in the untreated ones (190). A combination of
these drugs is being evaluated to treat MERS in
humans (MIRACLE trial) (191). These two protease
inhibitors (lopinavir and ritonavir), in combination
with ribavirin, gave encouraging clinical outcomes in
SARS patients, suggesting their therapeutic values
(165). However, in the current scenario, due to the
lack of specific therapeutic agents against SARS-
CoV-2, hospitalized patients confirmed for the
disease are given supportive care, like oxygen and
fluid therapy, along with antibiotic therapy for
managing secondary bacterial infections (192).
Patients with novel coronavirus or COVID-19
pneumonia who are mechanically ventilated often
require sedatives. analgesics, and even muscle
require sedatives, analgesics, and even muscle
relaxation drugs to prevent ventilator-related lung
injury associated with human-machine
incoordination (122). The result obtained from a
clinical study of four patients infected with COVID-
19 claimed that combination therapy using
lopinavir/ritonavir, arbidol, and Shufeng Jiedu
capsules (traditional Chinese medicine) was found to
be effective in managing COVID-19 pneumonia
(193). It is difficult to evaluate the therapeutic
potential of a drug or a combination of drugs for
managing a disease based on such a limited sample
size. Before choosing the ideal therapeutic agent for
the management of COVID-19, randomized clinical
control studies should be performed with a sufficient
study population.
Antiviral Drugs
Several classes of routinely used antiviral drugs,
like oseltamivir (neuraminidase inhibitor), acyclovir,
ganciclovir, and ribavirin, do not have any effect on
COVID-19 and, hence, are not recommended (187).
Oseltamivir, a neuraminidase inhibitor, has been
explored in Chinese hospitals for treating suspected
COVID-19 cases, although proven efficacy against
SARS-CoV-2 is still lacking for this drug (7). The in
vitro antiviral potential of FAD-approved drugs, viz.,
vitro antiviral potential of FAD-approved drugs, viz.,
ribavirin, penciclovir, nitazoxanide, nafamostat, and
chloroquine, tested in comparison to remdesivir and
favipiravir (broad-spectrum antiviral drugs) revealed
remdesivir and chloroquine to be highly effective
against SARS-CoV-2 infection in vitro (194).
Ribavirin, penciclovir, and favipiravir might not
possess noteworthy in vivo antiviral actions for
SARS-CoV-2, since higher concentrations of these
nucleoside analogs are needed in vitro to lessen the
viral infection. Both remdesivir and chloroquine are
being used in humans to treat other diseases, and
such safer drugs can be explored for assessing their
effectiveness in COVID-19 patients.
Several therapeutic agents, such as
lopinavir/ritonavir, chloroquine, and
hydroxychloroquine, have been proposed for the
clinical management of COVID-19 (299). A
molecular docking study, conducted in the RNA-
dependent RNA polymerase (RdRp) of SARS-CoV-2
using different commercially available
antipolymerase drugs, identified that drugs such as
ribavirin, remdesivir, galidesivir, tenofovir, and
sofosbuvir bind RdRp tightly, indicating their vast
potential to be used against COVID-19 (305). A
broad-spectrum antiviral drug that was developed in
the United States, tilorone dihydrochloride (tilorone),
the United States, tilorone dihydrochloride (tilorone),
was previously found to possess potent antiviral activity
against MERS, Marburg, Ebola, and
Chikungunya viruses (306). Even though it had
broad-spectrum activity, it was neglected for an
extended period. Tilorone is another antiviral drug
that might have activity against SARS-CoV-2.
Remdesivir, a novel nucleotide analog prodrug,
was developed for treating Ebola virus disease
(EVD), and it was also found to inhibit the
replication of SARS-CoV and MERS-CoV in
primary human airway epithelial cell culture systems
(195). Recently, in vitro study has proven that
remdesivir has better antiviral activity than lopinavir
and ritonavir. Further, in vivo studies conducted in
mice also identified that treatment with remdesivir
improved pulmonary function and reduced viral
loads and lung pathology both in prophylactic and
therapeutic regimens compared to
lopinavir/ritonavir-IFN-y treatment in MERS-CoV
infection (8). Remdesivir also inhibits a diverse
range of coronaviruses, including circulating human
COV, zoonotic bat CoV, and prepandemic zoonotic
CoV (195). Remdesivir is also considered the only
therapeutic drug that significantly reduces
pulmonary pathology (8). All these findings indicate
that remdesivir has to be further evaluated for its
that remdesivir has to be further evaluated for its
efficacy in the treatment of COVID-19 infection in
humans. The broad-spectrum activity exhibited by
remdesivir will help control the spread of disease in
the event of a new coronavirus outbreak.
Chloroquine is an antimalarial drug known to
possess antiviral activity due to its ability to block
virus-cell fusion by raising the endosomal pH
necessary for fusion. It also interferes with virus-
receptor binding by interfering with the terminal
glycosylation of SARS-CoV cellular receptors, such
as ACE2 (196). In a recent multicenter clinical trial
that was conducted in China, chloroquine phosphate
was found to exhibit both efficacy and safety in the
therapeutic management of SARS-CoV-2-associated
pneumonia (197). This drug is already included in
the treatment guidelines issued by the National
Health Commission of the People's Republic of
China. The preliminary clinical trials using
hydroxychloroquine, another aminoquinoline drug,
gave promising results. The COVID-19 patients
received 600 mg of hydroxychloroquine daily along
with azithromycin as a single-arm protocol. This
protocol was found to be associated with a
noteworthy reduction in viral load. Finally, it
resulted in a complete cure (271); however, the study
comprised a small population and, hence, the
comprised a small population and hence, the
possibility of misinterpretation could arise. However,
in another case study, the authors raised concerns
over the efficacy of hydroxychloroquine-
azithromycin in the treatment of COVID-19 patients,
since no observable effect was seen when they were
used. In some cases, the treatment was discontinued
due to the prolongation of the QT interval (307).
Hence, further randomized clinical trials are required
before concluding this matter.
Recently, another FDA-approved drug,
ivermectin, was reported to inhibit the in vitro
replication of SARS-CoV-2. The findings from this
study indicate that a single treatment of this drug was
able to induce an -5,000-fold reduction in the viral
RNA at 48 h in cell culture. (308). One of the main
disadvantages that limit the clinical utility of
ivermectin is its potential to cause cytotoxicity.
However, altering the vehicles used in the
formulations, the pharmacokinetic properties can be
modified, thereby having significant control over the
systemic concentration of ivermectin (338). Based
on the pharmacokinetic simulation, it was also found
that ivermectin may have limited therapeutic utility
in managing COVID-19, since the inhibitory
concentration that has to be achieved for effective
anti-SARS-CoV-2 activity is far higher than the
anti-SARS-CoV-2 activity is far higher than the
maximum plasma concentration achieved by
administering the approved dose (340). However,
ivermectin, being a host-directed agent, exhibits
antiviral activity by targeting a critical cellular
process of the mammalian cell. Therefore, the
administration of ivermectin, even at lower doses,
will reduce the viral load at a minor level. This slight
decrease will provide a great advantage to the
immune system for mounting a large-scale antiviral
response against SARS-CoV-2 (341). Further, a
combination of ivermectin and hydroxychloroquine
might have a synergistic effect, since ivermectin
reduces viral replication, while hydroxychloroquine
inhibits the entry of the virus in the host cell (339).
Further, in vivo studies and randomized clinical
control trials are required to understand the
mechanism as well as the clinical utility of this
promising drug.
Nafamostat is a potent inhibitor of MERS-CoV
that acts by preventing membrane fusion.
Nevertheless, it does not have any sort of inhibitory
action against SARS-CoV-2 infection (194).
Recently, several newly synthesized halogenated
triazole compounds were evaluated, using
fluorescence resonance energy transfer (FRET)-
based helicase assays, for their ability to inhibit
helicase activity.
Among the evaluated compounds, 4-(cyclopent-
1-en-3-ylamino)-5-[2-(4-
iodophenyl)hydrazinyl]-4H-1,2,4-triazole-3-thiol and
4-(cyclopent-1-en-3-ylamino)-5-[2-(4-
chlorophenyl)hydrazinyl]-4H-1,2,4-triazole-3-thiol
were found to be the most potent. These compounds
were used for in silico studies, and molecular
docking was accomplished into the active binding
site of MERS-CoV helicase nsp13 (21). Further
studies are required for evaluating the therapeutic
potential of these newly identified compounds in the
management of COVID-19 infection.
Passive Immunization/Antibody Therapy/MAb
Monoclonal antibodies (MAbs) may be helpful in
the intervention of disease in CoV-exposed
individuals. Patients recovering from SARS showed
robust neutralizing antibodies against this CoV
infection (164). A set of MAbs aimed at the MERS-
CoV S protein-specific domains, comprising six
specific epitope groups interacting with receptor-
binding, membrane fusion, and sialic acid-binding
sites, make up crucial entry tasks of S protein (198,
199). Passive immunization employing weaker and
strongly neutralizing antibodies provided
considerable protection in mice against a MERS
considerable protection in mice against a MERS-
CoV lethal challenge. Such antibodies may play a
crucial role in enhancing protective humoral
responses against the emerging CoVs by aiming
appropriate epitopes and functions of the S protein.
The cross-neutralization ability of SARS-CoV RBD-
specific neutralizing MAbs considerably relies on
the resemblance between their RBDs; therefore,
SARS-CoV RBD-specific antibodies could cross-
neutralized SL CoVs, i.e., bat-SL-CoV strain WIV1
(RBD with eight amino acid differences from SARS-
CoV) but not bat-SL-CoV strain SHC014 (24 amino
acid differences) (200).
Appropriate RBD-specific MAbs can be
recognized by a relative analysis of RBD of SARS-
CoV-2 to that of SARS-CoV, and cross-neutralizing
SARS-CoV RBD-specific MAbs could be explored
for their effectiveness against COVID-19 and further
need to be assessed clinically. The U.S.
biotechnology company Regeneron is attempting to
recognize potent and specific MAbs to combat
COVID-19. An ideal therapeutic option suggested
for SARS-CoV-2 (COVID-19) is the combination
therapy comprised of MAbs and the drug remdesivir
(COVID-19) (201). The SARS-CoV-specific human
MAL CR3022 is found to bind with SARS-CoV-2
RBD, indicating its potential as a therapeutic agent
RBD, indicating its potential as a therapeutic agent
in the management of COVID-19. It can be used
alone or in combination with other effective
neutralizing antibodies for the treatment and
prevention of COVID-19 (202). Furthermore, SARS-
CoV-specific neutralizing antibodies, like m396 and
CR3014, failed to bind the S protein of SARS-CoV-
2, indicating that a particular level of similarity is
mandatory between the RBDs of SARS-CoV and
SARS-CoV-2 for the cross-reactivity to occur.
Further assessment is necessary before
confirming the effectiveness of such combination
therapy. In addition, to prevent further community
and nosocomial spread of COVID-19, the
postprocedure risk management program should not
be neglected (309). Development of broad-spectrum
inhibitors against the human coronaviral pathogens
will help to facilitate clinical trials on the
effectiveness of such inhibitors against endemic and
emerging coronaviruses (203). A promising animal
study revealed the protective effect of passive
immunotherapy with immune serum from MERS-
immune camels on mice infected with MERS-CoV
(204). Passive immunotherapy using convalescent
plasma is another strategy that can be used for
treating COVID-19-infected, critically ill patients
(205).
The exploration of fully human antibodies
(human single-chain antibodies; HuscFvs) or
humanized nanobodies (single-domain antibodies;
sdAb, VH/VHH) could aid in blocking virus
replication, as these agents can traverse the virus-
infected cell membranes (transbodies) and can
interfere with the biological characteristics of the
replicating virus proteins. Such examples include
transbodies to the influenza virus, hepatitis C virus,
Ebola virus, and dengue virus (206). Producing
similar transbodies against intracellular proteins of
coronaviruses, such as papain-like proteases (PLpro),
cysteine-like protease (3CL pro), or other nsps, which
are essential for replication and transcription of the
virus, might formulate a practical move forward for a
safer and potent passive immunization approach for
virus-exposed persons and rendering therapy to
infected patients.
In a case study on five grimly sick patients
having symptoms of severe pneumonia due to
COVID-19, convalescent plasma administration was
found to be helpful in patients recovering
successfully. The convalescent plasma containing a
SARS-CoV-2-specific ELISA (serum) antibody titer
higher than 1:1,000 and neutralizing antibody titer
more significant than 40 was collected from the
recovered patients and used for plasma transfusion
recovered patients and used for plasma transfusion
twice in a volume of 200 to 250 ml on the day of
collection (310). At present, treatment for sepsis and
ARDS mainly involves antimicrobial therapy, source
control, and supportive care. Hence, the use of
therapeutic plasma exchange can be considered an
option in managing such severe conditions. Further
randomized trials can be designed to investigate its
efficacy (311).
Potential Therapeutic Agents
Potent therapeutics to combat SARS-CoV-2
infection include virus binding molecules, molecules
or inhibitors targeting particular enzymes implicated
in replication and transcription process of the virus,
helicase inhibitors, vital viral proteases and proteins,
protease inhibitors of host cells, endocytosis
inhibitors, short interfering RNA (siRNA),
neutralizing antibodies, MAbs against the host
receptor, MAbs interfering with the Si RBD,
antiviral peptide aimed at S2, and natural
drugs/medicines (7, 166, 186). The S protein acts as
the critical target for developing CoV antivirals, like
inhibitors of S protein and S cleavage, neutralizing
antibodies, RBD-ACE2 blockers, siRNAs, blockers
of the fusion core, and proteases (168).
All of these therapeutic approaches have revealed
All of these therapeutic approaches have revealed
both in vitro and in vivo anti-CoV potential.
Although in vitro research carried out with these
therapeutics showed efficacy, most need appropriate
support from randomized animal or human trials.
Therefore, they might be of limited applicability and
require trials against SARS-CoV-2 to gain practical
usefulness. The binding of SARS-CoV-2 with ACE2
leads to the exacerbation of pneumonia as a
consequence of the imbalance in the renin-
angiotensin system (RAS). The virus-induced
pulmonary inflammatory responses may be reduced
by the administration of ACE inhibitors (ACEI) and
angiotensin type-1 receptor (AT1R) (207).
Several investigations have suggested the use of
small-molecule inhibitors for the potential control of
SARS-CoV infections. Drugs of the FDA-approved
compound library were screened to identify four
small-molecule inhibitors of MERS-CoV
(chlorpromazine, chloroquine, loperamide, and
lopinavir) that inhibited viral replication. These
compounds also hinder SARS-CoV and human
CoVs (208). Therapeutic strategies involving the use
of specific antibodies or compounds that neutralize
cytokines and their receptors will help to restrain the
host inflammatory responses. Such drugs acting
specifically in the respiratory tract will help to
specifically in the respiratory tract will help to
reduce virus-triggered immune pathologies in
COVID-19 (209). The later stages of coronavirus-
induced inflammatory cascades are characterized by
the release of proinflammatory interleukin-1 (IL-1)
family members, such as IL-1 and IL-33. Hence,
there exists a possibility that the inflammation
associated with coronavirus can be inhibited by
utilizing anti-inflammatory cytokines that belong to
the IL-1 family (92). It has also been suggested that
the actin protein is the host factor that is involved in
cell entry and pathogenesis of SARS-CoV-2. Hence,
those drugs that modulate the biological activity of
this protein, like ibuprofen, might have some
therapeutic application in managing the disease
(174). The plasma angiotensin 2 level was found to
be markedly elevated in COVID-19 infection and
was correlated with viral load and lung injury.
Hence, drugs that block angiotensin receptors may
have potential for treating COVID-19 infection
(121). A scientist from Germany, named Rolf
Hilgenfeld, has been working on the identification of
drugs for the treatment of coronaviral infection since
the time of the first SARS outbreak (19).
The SARS-CoV S2 subunit has a significant
function in mediating virus fusion that provides entry
into the host cell. Heptad repeat 1 (HR1) and heptad
into the host cell. Heptad repeat 1 (HR1) and heptad
repeat 2 (HR2) can interact and form a six-helix
bundle that brings the viral and cellular membranes
in close proximity, facilitating its fusion. The
sequence alignment study conducted between
COVID-19 and SARS-CoV identified that the S2
subunits are highly conserved in these CoVs. The
HR1 and HR2 domains showed 92.6% and 100%
overall identity, respectively (210). From these
findings, we can confirm the significance of
COVID-19 HR1 and HR2 and their vital role in host
cell entry. Hence, fusion inhibitors target the HR1
domain of S protein, thereby preventing viral fusion
and entry into the host cell. This is another potential
therapeutic strategy that can be used in the
management of COVID-19. Other than the specific
therapy directed against COVID-19, general
treatments play a vital role in the enhancement of
host immune responses against the viral agent.
Inadequate nutrition is linked to the weakening of
the host immune response, making the individual
more susceptible. The role played by nutrition in
disease susceptibility should be measured by
evaluating the nutritional status of patients with
COVID-19 (205).
Animal Models and Cell Cultures
For evaluating the potential of vaccines and
therapeutics against CoVs, including SARS-CoV,
MERS-CoVs, and the presently emerging SARS-
COV-2, suitable animal models that can mimic the
clinical disease are needed (211, 212). Various
animal models were assessed for SARS- and MERS-
CoVs, such as mice, guinea pigs, golden Syrian
hamsters, ferrets, rabbits, nonhuman primates like
rhesus macaques and marmosets, and cats (185,
213–218). The specificity of the virus to hACE2
(receptor of SARS-CoV) was found to be a
significant barrier in developing animal models.
Consequently, a SARS-CoV transgenic mouse model
has been developed by inserting the hACE2 gene
into the mouse genome (219). The inability of
MERS-CoV to replicate in the respiratory tracts of
animals (mice, hamsters, and ferrets) is another
limiting factor. However, with genetic engineering, a
288-330++ MERS-CoV genetically modified mouse
model was developed and now is in use for the
assessment of novel drugs and vaccines against
MERS-CoV (220). In the past, small animals (mice
or hamsters) have been targeted for being closer to a
humanized structure, such as mouse DPP4 altered
with human DPP4 (HDPP4), HDPP4-transduced
mice, and hDPP4-Tg mice (transgenic for expressing
mice, and hDPP4-Tg mice (transgenic for expressing
hDPP4) for MERS-CoV infection (221). The
CRISPR-Cas9 gene-editing tool has been used for
inserting genomic alterations in mice, making them
susceptible to MERS-CoV infection (222). Efforts
are under way to recognize suitable animal models
for SARS-CoV2/COVID-19, identify the receptor
affinity of this virus, study pathology in experimental
animal models, and explore virus-specific immune
responses and protection studies, which together
would increase the pace of efforts being made for
developing potent vaccines and drugs to counter this
emerging virus. Cell lines, such as monkey epithelial
cell lines (LLC-MK2 and Vero-B4), goat lung cells,
alpaca kidney cells, dromedary umbilical cord cells,
and advanced ex vivo three-dimensional
tracheobronchial tissue, have been explored to study
human CoVs (MERS-CoV) (223, 224). Vero and
Huh-7 cells (human liver cancer cells) have been
used for isolating SARS-CoV-2 (194).
Recently, an experimental study with rhesus
monkeys as animal models revealed the absence of
any viral loads in nasopharyngeal and anal swabs,
and no viral replication was recorded in the primary
tissues at a time interval of 5 days post-reinfection in
reexposed monkeys (274). The subsequent
virological, radiological, and pathological
virological, radiological and pathological
observations indicated that the monkeys with
reexposure had no recurrence of COVID-19, like the
SARS-CoV-2-infected monkeys without rechallenge.
These findings suggest that primary infection with
SARS-CoV-2 could protect from later exposures to
the virus, which could help in defining disease
prognosis and crucial inferences for designing and
developing potent vaccines against COVID-19
(274).
PREVENTION, CONTROL, AND
MANAGEMENT
In contrast to their response to the 2002 SARS
outbreak, China has shown immense political
openness in reporting the COVID-19 outbreak
promptly. They have also performed rapid
sequencing of COVID-19 at multiple levels and
shared the findings globally within days of
identifying the novel virus (225). The move made by
China opened a new chapter in global health security
and diplomacy. Even though complete lockdown was
declared following the COVID-19 outbreak in
Wuhan, the large-scale movement of people has
resulted in a radiating spread of infections in the
surrounding provinces as well as to several other
countries. Large-scale screening programs might
countries. Large-scale screening programs might
help us to control the spread of this virus. However,
this is both challenging as well as time-consuming
due to the present extent of infection (226). The
current scenario demands effective implementation
of vigorous prevention and control strategies owing
to the prospect of COVID-19 for nosocomial
infections (68). Follow-ups of infected patients by
telephone on day 7 and day 14 are advised to avoid
any further unintentional spread or nosocomial
transmission (312). The availability of public data
sets provided by independent analytical teams will
act as robust evidence that would guide us in
designing interventions against the COVID-19
outbreak. Newspaper reports and social media can be
used to analyze and reconstruct the progression of an
outbreak. They can help us to obtain detailed patient-
level data in the early stages of an outbreak (227).
Immediate travel restrictions imposed by several
countries might have contributed significantly to
preventing the spread of SARS-CoV-2 globally (89,
228). Following the outbreak, a temporary ban was
imposed on the wildlife trade, keeping in mind the
possible role played by wild animal species in the
origin of SARS-CoV-2/COVID-19 (147). Making a
permanent and bold decision on the trade of wild
animal species is necessary to prevent the possibility
animal species is necessary to prevent the possibility
of virus spread and initiation of an outbreak due to
zoonotic spillover (1).
Personal protective equipment (PPE), like face
masks, will help to prevent the spread of respiratory
infections like COVID-19. Face masks not only
protect from infectious aerosols but also prevent the
transmission of disease to other susceptible
individuals while traveling through public transport
systems (313). Another critical practice that can
reduce the transmission of respiratory diseases is the
maintenance of hand hygiene. However, the efficacy
of this practice in reducing the transmission of
respiratory viruses like SARS-CoV-2 is much
dependent upon the size of droplets produced. Hand
hygiene will reduce disease transmission only if the
virus is transmitted through the formation of large
droplets (314). Hence, it is better not to
overemphasize that hand hygiene will prevent the
transmission of SARS-CoV-2, since it may produce a
false sense of safety among the general public that
further contributes to the spread of COVID-19. Even
though airborne spread has not been reported in
SARS-CoV-2 infection, transmission can occur
through droplets and fomites, especially when there
is close, unprotected contact between infected and
susceptible individuals. Hence, hand hygiene is
health emergency on 31 January 2020; subsequently,
on 11 March 2020, they declared it a pandemic
situation. At present, we are not in a position to
effectively treat COVID-19, since neither approved
vaccines nor specific antiviral drugs for treating
human CoV infections are available (7–9). Most
nations are currently making efforts to prevent the
further spreading of this potentially deadly virus by
implementing preventive and control strategies.
In domestic animals, infections with CoVs are
associated with a broad spectrum of pathological
conditions. Apart from infectious bronchitis virus,
canine respiratory CoV, and mouse hepatitis virus,
CoVs are predominantly associated with
gastrointestinal diseases (10). The emergence of
novel CoVs may have become possible because of
multiple CoVs being maintained in their natural host,
which could have favored the probability of genetic
recombination (10). High genetic diversity and the
ability to infect multiple host species are a result of
high-frequency mutations in CoVs, which occur due
to the instability of RNA-dependent RNA
polymerases along with higher rates of homologous
RNA recombination (10, 11). Identifying the origin
of SARS-CoV-2 and the pathogen’s evolution will be
helpful for disease surveillance (12), development of
new targeted drugs, and prevention of further
epidemics (13). The most common symptoms
associated with COVID-19 are fever, cough,
dyspnea, expectoration, headache, and myalgia or
fatigue.
In contrast, less common signs at the time of
hospital admission include diarrhea, hemoptysis, and
shortness of breath (14). Recently, individuals with
asymptomatic infections were also suspected of
transmitting infections, which further adds to the
complexity of disease transmission dynamics in
COVID-19 infections (1). Such efficient responses
require in-depth knowledge regarding the virus,
which currently is a novel agent; consequently,
further studies are required.
Comparing the genome of SARS-CoV-2 with that
of the closely related SARS/SARS-like COV
revealed that the sequence coding for the spike
protein, with a total length of 1,273 amino acids,
showed 27 amino acid substitutions. Six of these
substitutions are in the region of the receptor-binding
domain (RBD), and another six substitutions are in
the underpinning subdomain (SD) (16). Phylogenetic
analyses have revealed that SARS-CoV-2 is closely
related (88% similarity) to two SARS-like COVs
derived from bat SARS-like COVs (bat-SL
CoVZC45 and bat-SL-COVZXC21) (Fig. 1).
Furthermore, SARS-CoV-2 is genetically distinct
from SARS-CoV (79% similarity) and MERS-CoV
(nearly 50%) (17). COVID-19 is associated with
afflictions of the lungs in all cases and generated
characteristic chest computer tomography findings,
such as the presence of multiple lesions in lung lobes
that appear as dense, ground-glass opaque structures
that occasionally coexist with consolidation shadows
(18).
Some therapeutic options for treating COVID-19
showed efficacy in in vitro studies; however, to date,
these treatments have not undergone any randomized
animal or human clinical trials, which limit their
practical applicability in the current pandemic (7, 9,
19-21).
The present comprehensive review describes the
various features of SARS-CoV-2/COVID-19 causing
the current disease outbreaks and advances in
diagnosis and developing vaccines and therapeutics.
It also provides a brief comparison with the earlier
SARS and MERS COVs, the veterinary perspective
of CoVs and this emerging novel pathogen, and an
evaluation of the zoonotic potential of similar CoVs
to provide feasible One Health strategies for the
management of this fatal virus (22–367).
THE VIRUS (SARS-CoV-2)
Coronaviruses are positive-sense RNA viruses
having an extensive and promiscuous range of
natural hosts and affect multiple systems (23, 24).
Coronaviruses can cause clinical diseases in humans
that may extend from the common cold to more
severe respiratory diseases like SARS and MERS
(17, 279). The recently emerging SARS-CoV-2 has
wrought havoc in China and caused a pandemic
situation in the worldwide nonulation leading to
wrought havoc in China and caused a pandemic
situation in the worldwide population, leading to
disease outbreaks that have not been controlled to
date, although extensive efforts are being put in
place to counter this virus (25). This virus has been
proposed to be designated/named severe acute
respiratory syndrome coronavirus 2 (SARS-CoV-2)
by the International Committee on Taxonomy of
Viruses (ICTV), which determined the virus belongs
to the Severe acute respiratory syndrome-related
coronavirus category and found this virus is related
to SARS-CoVs (26). SARS-CoV-2 is a member of
the order Nidovirales, family Coronaviridae,
subfamily Orthocoronavirinae, which is subdivided
into four genera, viz., Alphacoronavirus,
Betacoronavirus, Gammacoronavirus, and
Deltacoronavirus (3, 27). The genera
Alphacoronavirus and Betacoronavirus originate
from bats, while Gammacoronavirus and
Deltacoronavirus have evolved from bird and swine
gene pools (24, 28, 29, 275).
Coronaviruses possess an unsegmented, single-
stranded, positive-sense RNA genome of around 30
kb, enclosed by a 5'-cap and 3'-poly(A) tail (30). The
genome of SARS-CoV-2 is 29,891 bp long, with a
G+C content of 38% (31). These viruses are
encircled with an envelope containing viral
encircled with an envelope containing viral
nucleocapsid. The nucleocapsids in CoVs are
arranged in helical symmetry, which reflects an
atypical attribute in positive-sense RNA viruses (30).
The electron micrographs of SARS-CoV-2 revealed
a diverging spherical outline with some degree of
pleomorphism, virion diameters varying from 60 to
140 nm, and distinct spikes of 9 to 12 nm, giving the
virus the appearance of a solar corona (3). The CoV
genome is arranged linearly as 5'-leader-UTR-
replicase-structural genes (S-E-M-N)-3' UTR-
poly(A) (32). Accessory genes, such as 3a/b, 4a/b,
and the hemagglutinin-esterase gene (HE), are also
seen intermingled with the structural genes (30).
SARS-CoV-2 has also been found to be arranged
similarly and encodes several accessory proteins,
although it lacks the HE, which is characteristic of
some betacoronaviruses (31). The positive-sense
genome of CoVs serves as the mRNA and is
translated to polyprotein la/lab (ppla/lab) (33). A
replication-transcription complex (RTC) is formed in
double-membrane vesicles (DMVs) by nonstructural
proteins (nsps), encoded by the polyprotein gene
(34). Subsequently, the RTC synthesizes a nested set
of subgenomic RNAs (sgRNAs) via discontinuous
transcription (35).
Based on molecular characterization, SARSC
oV-2 is considered a new Betacoronavirus
belonging to the subgenus Sarbecovirus (3). A few
other critical zoonotic viruses (MERS-related CoV
and SARS-related CoV) belong to the same genus.
However, SARS-CoV-2 was identified as a distinct
virus based on the percent identity with other
Betacoronavirus; conserved open reading frame 1a/b
(ORFla/b) is below 90% identity (3). An overall
80% nucleotide identity was observed between
SARS-CoV-2 and the original SARS-CoV, along
with 89% identity with ZC45 and ZXC21 SARS
related CoVs of bats (2, 31, 36). In addition, 82%
identity has been observed between SARS-CoV-2
and human SARS-CoV Tor2 and human SARS-CoV
BJ01 2003 (31). A sequence identity of only 51.8%
was observed between MERS-related CoV and the
recently emerged SARS-CoV-2 (37). Phylogenetic
analysis of the structural genes also revealed that
SARS-CoV-2 is closer to bat SARS-related CoV.
Therefore, SARS-CoV-2 might have originated from
bats, while other amplifier hosts might have played a
role in disease transmission to humans (31). Of note,
the other two zoonotic CoVs (MERS-related CoV
and SARS-related CoV) also originated from bats
(38, 39). Nevertheless, for SARS and MERS, civet
(30, 59). Nevertheless, TOT SARS and MERS, Civet
cat and camels, respectively, act as amplifier hosts
(40, 41).
Coronavirus genomes and subgenomes encode
six ORFs (31). The majority of the 5' end is occupied
by ORF1a/b, which produces 16 nsps. The two
polyproteins, ppla and pplab, are initially produced
from ORF1a/b by a -1 frameshift between ORFla
and ORF16 (32). The virus-encoded proteases cleave
polyproteins into individual nsps (main protease
[Mpro), chymotrypsin-like protease [3CL pro], and
papain-like proteases [PLPs]) (42). SARS-CoV-2
also encodes these nsps, and their functions have
been elucidated recently (31). Remarkably, a
difference between SARS-CoV-2 and other CoVs is
the identification of a novel short putative protein
within the ORF3 band, a secreted protein with an
alpha helix and beta-sheet with six strands encoded
by ORF8 (31).
Coronaviruses encode four major structural
proteins, namely, spike (S), membrane (M), envelope
(E), and nucleocapsid (N), which are described in
detail below.
S Glycoprotein
Coronavirus S protein is a large, multifunctional
class I viral transmembrane protein. The size of this
Coronavirus S protein is a large, multifunctional
class I viral transmembrane protein. The size of this
abundant S protein varies from 1,160 amino acids
(IBV, infectious bronchitis virus, in poultry) to 1,400
amino acids (FCOV, feline coronavirus) (43). It lies
in a trimer on the virion surface, giving the virion a
corona or crown-like appearance. Functionally it is
required for the entry of the infectious virion
particles into the cell through interaction with
various host cellular receptors (44).
Furthermore, it acts as a critical factor for tissue
tropism and the determination of host range (45).
Notably, S protein is one of the vital
immunodominant proteins of COVs capable of
inducing host immune responses (45). The
ectodomains in all CoVs S proteins have similar
domain organizations, divided into two subunits, S1
and S2 (43). The first one, S1, helps in host receptor
binding, while the second one, S2, accounts for
fusion. The former (S1) is further divided into two
subdomains, namely, the N-terminal domain (NTD)
and C-terminal domain (CTD). Both of these
subdomains act as receptor-binding domains,
interacting efficiently with various host receptors
(45). The S1 CTD contains the receptor-binding
motif (RBM). In each coronavirus spike protein, the
trimeric S1 locates itself on top of the trimeric S2
trimeric Si locates itself on top of the trimeric S2
stalk (45). Recently, structural analyses of the S
proteins of COVID-19 have revealed 27 amino acid
substitutions within a 1,273-amino-acid stretch (16).
Six substitutions are located in the RBD (amino
acids 357 to 528), while four substitutions are in the
RBM at the CTD of the S1 domain (16). Of note, no
amino acid change is seen in the RBM, which binds
directly to the angiotensin-converting enzyme-2
(ACE2) receptor in SARS-CoV (16, 46). At present,
the main emphasis is knowing how many differences
would be required to change the host tropism.
Sequence comparison revealed 17 nonsynonymous
changes between the early sequence of SARS-CoV-2
and the later isolates of SARS-CoV. The changes
were found scattered over the genome of the virus,
with nine substitutions in ORFlab, ORF8 (4
substitutions), the spike gene (3 substitutions), and
ORF7a (single substitution) (4). Notably, the same
nonsynonymous changes were found in a familial
cluster, indicating that the viral evolution happened
during person-to-person transmission (4, 47). Such
adaptive evolution events are frequent and constitute
a constantly ongoing process once the virus spreads
among new hosts (47). Even though no functional
changes occur in the virus associated with this
adaptive evolution, close monitoring of the viral
adaptive evolution, close monitoring of the viral
mutations that occur during subsequent human-to-
human transmission is warranted.
M Protein
The M protein is the most abundant viral protein
present in the virion particle, giving a definite shape
to the viral envelope (48). It binds to the
nucleocapsid and acts as a central organizer of
coronavirus assembly (49). Coronavirus M proteins
are highly diverse in amino acid contents but
maintain overall structural similarity within different
genera (50). The M protein has three transmembrane
domains, flanked by a short amino terminus outside
the virion and a long carboxy terminus inside the
virion (50). Overall, the viral scaffold is maintained
by M-M interaction. Of note, the M protein of
SARS-CoV-2 does not have an amino acid
substitution compared to that of SARS-CoV (16).
E Protein
The coronavirus E protein is the most enigmatic
and smallest of the major structural proteins (51). It
plays a multifunctional role in the pathogenesis,
assembly, and release of the virus (52). It is a small
integral membrane polypeptide that acts as a
viroporin (ion channel) (53). The inactivation or
absence of this protein is related to the altered
virulence of coronaviruses due to changes in
morphology and tropism (54). The E protein consists
of three domains, namely, a short hydrophilic amino
terminal, a large hydrophobic transmembrane
domain, and an efficient C-terminal domain (51).
The SARS-CoV-2 E protein reveals a similar amino
acid constitution without any substitution (16).
N Protein
The N protein of coronavirus is multipurpose.
Among several functions, it plays a role in complex
formation with the viral genome, facilitates M
protein interaction needed during virion assembly,
and enhances the transcription efficiency of the virus
(55, 56). It contains three highly conserved and
distinct domains, namely, an NTD, an RNA-binding
domain or a linker region (LKR), and a CTD (57).
The NTD binds with the 3' end of the viral genome,
perhaps via electrostatic interactions, and is highly
diverged both in length and sequence (58). The
charged LKR is serine and arginine rich and is also
known as the SR (serine and arginine) domain (59).
The LKR is capable of direct interaction with in vitro
RNA interaction and is responsible for cell signaling
(60, 61).It also modulates the antiviral response of
the host by working as an antagonist for interferon
N Protein
The N protein of coronavirus is multipurpose.
Among several functions, it plays a role in complex
formation with the viral genome, facilitates M
protein interaction needed during virion assembly,
and enhances the transcription efficiency of the virus
(55, 56). It contains three highly conserved and
distinct domains, namely, an NTD, an RNA-binding
domain or a linker region (LKR), and a CTD (57).
The NTD binds with the 3' end of the viral genome,
perhaps via electrostatic interactions, and is highly
diverged both in length and sequence (58). The
charged LKR is serine and arginine rich and is also
known as the SR (serine and arginine) domain (59).
The LKR is capable of direct interaction with in vitro
RNA interaction and is responsible for cell signaling
(60, 61). It also modulates the antiviral response of
the host by working as an antagonist for interferon
(IFN) and RNA interference (62). Compared to that
of SARS-CoV, the N protein of SARS-CoV-2
possess five amino acid mutations, where two are in
the intrinsically dispersed region (IDR; positions 25
and 26), one each in the NTD (position 103), LKR
(position 217), and CTD (position 334) (16).
nsps and Accessory Proteins
nsps and Accessory Proteins
Besides the important structural proteins, the
SARS-CoV-2 genome contains 15 nsps, nspl to
nsp10 and nsp12 to nsp16, and 8 accessory proteins
(3a, 3b, p6, 7a, 7b, 8b, 9b, and ORF14) (16). All
these proteins play a specific role in viral replication
(27). Unlike the accessory proteins of SARS-CoV,
SARS-CoV-2 does not contain 8a protein and has a
longer 8b and shorter 3b protein (16). The nsp7,
nsp13, envelope, matrix, and p6 and 8b accessory
proteins have not been detected with any amino acid
substitutions compared to the sequences of other
coronaviruses (16).
The virus structure of SARS-CoV-2 is depicted in
Fig. 2.

We assessed the nucleotide percent similarity
using the MegAlign software program, where the
similarity between the novel SARS-CoV-2 isolates
was in the range of 99.4% to 100%. Among the other
Serbecovirus CoV sequences, the novel SARS-CoV-
2 sequences revealed the highest similarity to bat-
SL-CoV, with nucleotide percent identity ranges
between 88.12 and 89.65%. Meanwhile, earlier
reported SARS-CoVs showed 70.6 to 74.9%
similarity to SARS-CoV-2 at the nucleotide level.
Further, the nucleotide percent similarity was 55.4%,
45.5% to 47.9%, 46.2% to 46.6%, and 45.0% to
46.3% to the other four subgenera, namely,
Hibecovirus, Nobecovirus, Merbecovirus, and
Embecovirus, respectively. The percent similarity
index of current outbreak isolates indicates a close
relationship between SARS-CoV-2 isolates and bat-
SL-CoV, indicating a common origin. However,
particular pieces of evidence based on further
complete genomic analysis of current isolates are
necessary to draw any conclusions, although it was
ascertained that the current novel SARS-CoV-2
isolates belong to the subgenus Sarbecovirus in the
diverse range of beta coronaviruses. Their possible
ancestor was hypothesized to be from bat CoV
strains, wherein bats might have played a crucial role
in harboring this class of viruses.
Splits Tree phylogeny analysis.
In the unrooted phylogenetic tree of different
betacoronaviruses based on the S protein, virus
sequences from different subgenera grouped into
separate clusters. SARS-CoV-2 sequences from
Wuhan and other countries exhibited a close
relationship and appeared in a single cluster (Fig. 1).
The CoVs from the subgenus Sarbecovirus appeared
jointly in SplitsTree and divided into three
subclusters, namely, SARS-CoV-2, bat-SARS-like-
CoV (bat-SL-CoV), and SARS-CoV (Fig. 1). In the
case of other subgenera, like Merbecovirus, all of the
sequences grouped in a single cluster, whereas in
Embecovirus, different species, comprised of canine
respiratory CoVs, bovine CoVs, equine CoVs, and
human CoV strain (OC43), grouped in a common
cluster. Isolates in the subgenera Nobecovorus and
Hibecovirus were found to be placed separately
away from other reported SARS-CoVs but shared a
bat origin.
CURRENT WORLDWIDE SCENARIO OF
SARS-CoV-2
This novel virus, SARS-CoV-2, comes under the
subgenus Sarbecovirus of the Orthocoronavirinae
subfamily and is entirely different from the viruses
responsible for MERS-CoV and SARS-CoV (3). The
newly emerged SARS-CoV-2 is a group 2B
coronavirus (2). The genome sequences of SARS-
CoV-2 obtained from patients share 79.5% sequence
similarity to the sequence of SARS-CoV (63).
As of 13 May 2020, a total of 4,170,424
confirmed cases of COVID-19 (with 287,399 deaths)
have been reported in more than 210 affected
countries worldwide (WHO Situation Report 114
Initially, the epicenter of the SARS-CoV-2
pandemic was China, which reported a significant
number of deaths associated with COVID-19, with
84,458 laboratory-confirmed cases and 4,644 deaths
as of 13 May 2020 (Fig. 4). As of 13 May 2020,
SARS-CoV-2 confirmed cases have been reported in
more than 210 countries apart from China (Fig. 3
and 4) (WHO Situation Report 114) (25, 64).
COVID-19 has been reported on all continents
except Antarctica. For many weeks, Italy was the
focus of concerns regarding the large number of
cases, with 221,216 cases and 30,911 deaths, but
now, the United States is the country with the largest
number of cases, 1,322,054, and 79,634 deaths.
Now, the United Kingdom has even more cases
(226,4671) and deaths (32,692) than Italy. A John
Hopkins University web platform has provided daily
updates on the basic epidemiology of the COVID-19
outbreak
COVID-19 has also been confirmed on a cruise
ship, named Diamond Princess, quarantined in
Japanese waters (Port of Yokohama), as well as on
other cruise ships around the world (239) (Fig. 3).
The significant events of the SARS-CoV-2/COVID-
19 virus outbreak occurring since 8 December 2019
are presented as a timeline in Fig. 5.
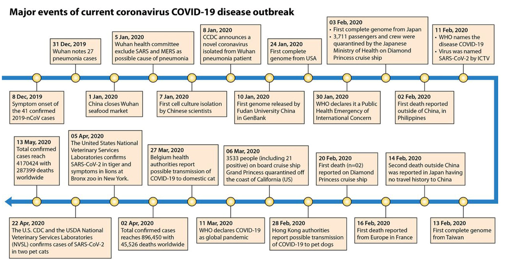
FIG 5 Timeline depicting the significant events that
occurred during the SARS-CoV-2/COVID-19 virus
outbreak. The timeline describes the significant events
during the current SARS-CoV-2 outbreak, from 8
December 2019 to 13 May 2020.
At the beginning, China experienced the majority
of the burden associated with COVID-19 in the form
of disease morbidity and mortality (65), but over
time the COVID-19 menace moved to Europe,
particularly Italy and Spain, and now the United
States has the highest number of confirmed cases
and deaths. The COVID-19 outbreak has also been
associated with severe economic impacts globally
due to the sudden interruption of global trade and
supply chains that forced multinational companies to
make decisions that led to significant economic
losses (66). The recent increase in the number of
confirmed critically ill patients with COVID-19 has
already surpassed the intensive care supplies, limiting
intensive care services to only a small
portion of critically ill patients (67). This might also
have contributed to the increased case fatality rate
observed in the COVID-19 outbreak.
Viewpoint on SARS-CoV-2 Transmission,
Spread, and Emergence
The novel coronavirus was identified within 1
month (28 days) of the outbreak. This is impressively
fast compared to the time taken to identify SARS-
CoV reported in Foshan, Guangdong Province,
China (125 days) (68). Immediately after the
confirmation of viral etiology, the Chinese
virologists rapidly released the genomic sequence of
SARS-CoV-2, which played a crucial role in
controlling the spread of this newly emerged novel
coronavirus to other parts of the world (69). The
possible origin of SARS-CoV-2 and the first mode of
possible origin of SARS-CO V-2 and the first mode of
disease transmission are not yet identified (70).
Analysis of the initial cluster of infections suggests
that the infected individuals had a common exposure
point, a seafood market in Wuhan, Hubei Province,
China (Fig. 6). The restaurants of this market are
well-known for providing different types of wild
animals for human consumption (71). The Huanan
South China Seafood Market also sells live animals,
such as poultry, bats, snakes, and marmots (72). This
might be the point where zoonotic (animal-to-
human transmission occurred (71). Although
SARS-CoV-2 is alleged to have originated from an
animal host (zoonotic origin) with further human-to-
human transmission (Fig. 6), the likelihood of
foodborne transmission should be ruled out with
further investigations, since it is a latent possibility
(1). Additionally, other potential and expected routes
would be associated with transmission, as in other
respiratory viruses, by direct contact, such as shaking
contaminated hands, or by direct contact with
contaminated surfaces (Fig. 6). Still, whether blood
transfusion and organ transplantation (276), as well
as transplacental and perinatal routes, are possible
routes for SARS-CoV-2 transmission needs to be
determined (Fig. 6).
From experience with several outbreaks
associated with known emerging viruses, higher
pathogenicity of a virus is often associated with
lower transmissibility. Compared to emerging
viruses like Ebola virus, avian H7N9, SARS-CoV,
and MERS-CoV, SARS-CoV-2 has relatively lower
pathogenicity and moderate transmissibility (15).
The risk of death among individuals infected with
COVID-19 was calculated using the infection
fatality risk (IFR). The IFR was found to be in the
range of 0.3% to 0.6%, which is comparable to that
of a previous Asian influenza pandemic (1957 to
1958) (73, 277).
Notably, the reanalysis of the COVID-19
pandemic curve from the initial cluster of cases
pointed to considerable human-to-human
transmission. It is opined that the exposure history of
SARS-CoV-2 at the Wuhan seafood market
originated from human-to-human transmission rather
than animal-to-human transmission (74); however, in
light of the zoonotic spillover in COVID-19, is too
early to fully endorse this idea (1). Following the
initial infection, human-to-human transmission has
been observed with a preliminary reproduction
number (Ro) estimate of 1.4 to 2.5 (70, 75), and
recently it is estimated to be 2.24 to 3.58 (76). In
another study, the average reproductive number of
another study, the average reproductive number of
COVID-19 was found to be 3.28, which is
significantly higher than the initial WHO estimate of
1.4 to 2.5 (77). It is too early to obtain the exact Ro
value, since there is a possibility of bias due to
insufficient data. The higher Ro value is indicative of
the more significant potential of SARS-CoV-2
transmission in a susceptible population. This is not
the first time where the culinary practices of China
have been blamed for the origin of novel coronavirus
infection in humans. Previously, the animals present
in the live-animal market were identified to be the
intermediate hosts of the SARS outbreak in China
(78). Several wildlife species were found to harbor
potentially evolving coronavirus strains that can
overcome the species barrier (79). One of the main
principles of Chinese food culture is that live-
slaughtered animals are considered more nutritious
(5).
After 4 months of struggle that lasted from
December 2019 to March 2020, the COVID-19
situation now seems under control in China. The wet
animal markets have reopened, and people have
started buying bats, dogs, cats, birds, scorpions,
badgers, rabbits, pangolins (scaly anteaters), minks,
soup from palm civet, ostriches, hamsters, snapping
turtles, ducks, fish, Siamese crocodiles, and other
turtles, ducks, fish, Siamese crocodiles, and other
animal meats without any fear of COVID-19. The
Chinese government is encouraging people to feel
they can return to normalcy. However, this could be
a risk, as it has been mentioned in advisories that
people should avoid contact with live-dead animals
as much as possible, as SARS-CoV-2 has shown
zoonotic spillover. Additionally, we cannot rule out
the possibility of new mutations in the same virus
being closely related to contact with both animals
and humans at the market (284). In January 2020,
China imposed a temporary ban on the sale of live-
dead animals in wet markets. However, now
hundreds of such wet markets have been reopened
without optimizing standard food safety and
sanitation practices (286).
With China being the most populated country in
the world and due to its domestic and international
food exportation policies, the whole world is now
facing the menace of COVID-19, including China
itself. Wet markets of live-dead animals do not
maintain strict food hygienic practices. Fresh blood
splashes are present everywhere, on the floor and
tabletops, and such food customs could encourage
many pathogens to adapt, mutate, and jump the
species barrier. As a result, the whole world is
suffering from novel SARS-CoV-2, with more than
suffering from novel SARS-CoV-2, with more than
4,170,424 cases and 287,399 deaths across the globe.
There is an urgent need for a rational international
campaign against the unhealthy food practices of
China to encourage the sellers to increase hygienic
food practices or close the crude live-dead animal
wet markets. There is a need to modify food policies
at national and international levels to avoid further
life threats and economic consequences from any
emerging or reemerging pandemic due to close
animal-human interaction (285).
Even though individuals of all ages and sexes are
susceptible to COVID-19, older people with an
underlying chronic disease are more likely to
become severely infected (80). Recently, individuals
with asymptomatic infection were also found to act
as a source of infection to susceptible individuals
(81). Both the asymptomatic and symptomatic
patients secrete similar viral loads, which indicates
that the transmission capacity of asymptomatic or
minimally symptomatic patients is very high. Thus,
SARS-CoV-2 transmission can happen early in the
course of infection (82). Atypical clinical
manifestations have also been reported in COVID-19
in which the only reporting symptom was fatigue.
Such patients may lack respiratory signs, such as
fever, cough, and sputum (83). Hence, the clinicians
fever, cough, and sputum (83). Hence, the clinicians
must be on the look-out for the possible occurrence
of atypical clinical manifestations to avoid the
possibility of missed diagnosis. The early
transmission ability of SARS-CoV-2 was found to be
similar to or slightly higher than that of SARS-CoV,
reflecting that it could be controlled despite
moderate to high transmissibility (84).
Increasing reports of SARS-CoV-2 in sewage and
wastewater warrants the need for further
investigation due to the possibility of fecal-oral
transmission. SARS-CoV-2 present in environmental
compartments such as soil and water will finally end
up in the wastewater and sewage sludge of treatment
plants (328). Therefore, we have to reevaluate the
current wastewater and sewage sludge treatment
procedures and introduce advanced techniques that
are specific and effective against SARS-CoV-2.
Since there is active shedding of SARS-CoV-2 in the
stool, the prevalence of infections in a large
population can be studied using wastewater-based
epidemiology. Recently, reverse transcription-
quantitative PCR (RT-qPCR) was used to enumerate
the copies of SARS-CoV-2 RNA concentrated from
wastewater collected from a wastewater treatment
plant (327). The calculated viral RNA copy numbers
determine the number of infected individuals. The
route warrants the introduction of negative fecal viral
nucleic acid test results as one of the additional
discharge criteria in laboratory-confirmed cases of
COVID-19 (326).
The COVID-19 pandemic does not have any
novel factors, other than the genetically unique
pathogen and a further possible reservoir. The cause
and the likely future outcome are just repetitions of
our previous interactions with fatal coronaviruses.
The only difference is the time of occurrence and the
genetic distinctness of the pathogen involved.
Mutations on the RBD of CoVs facilitated their
capability of infecting newer hosts, thereby
expanding their reach to all corners of the world
(85). This is a potential threat to the health of both
animals and humans. Advanced studies using
Bayesian phylogeographic reconstruction identified
the most probable origin of SARS-CoV-2 as the bat
SARS-like coronavirus, circulating in the
Rhinolophus bat family (86).
Phylogenetic analysis of 10 whole-genome
sequences of SARS-CoV-2 showed that they are
related to two CoVs of bat origin, namely, bat-SL-
CoVZC45 and bat-SL-COVZXC21, which were
reported during 2018 in China (17). It was reported
that SARS-CoV-2 had been confirmed to use ACE2
as an entry receptor while exhibiting an RBD similar
as an entry receptor while exhibiting an RBD similar
to that of SARS-CoV (17, 87, 254, 255). Several
countries have provided recommendations to their
people traveling to China (88, 89). Compared to the
previous coronavirus outbreaks caused by SARS-
CoV and MERS-CoV, the efficiency of SARS-CoV-
2 human-to-human transmission was thought to be
less. This assumption was based on the finding that
health workers were affected less than they were in
previous outbreaks of fatal coronaviruses (2).
Superspreading events are considered the main
culprit for the extensive transmission of SARS and
MERS (90, 91). Almost half of the MERS-CoV
cases reported in Saudi Arabia are of secondary
origin that occurred through contact with infected
asymptomatic or symptomatic individuals through
human-to-human transmission (92). The occurrence
of superspreading events in the COVID-19 outbreak
cannot be ruled out until its possibility is evaluated.
Like SARS and MERS, COVID-19 can also infect
the lower respiratory tract, with milder symptoms
(27). The basic reproduction number of COVID-19
has been found to be in the range of 2.8 to 3.3 based
on real-time reports and 3.2 to 3.9 based on predicted
infected cases (84).
Coronaviruses in Humans—SARS, MERS,
and COVID-19
Coronavirus infection in humans is commonly
associated with mild to severe respiratory diseases,
with high fever, severe inflammation, cough, and
internal organ dysfunction that can even lead to
death (92). Most of the identified coronaviruses
cause the common cold in humans. However, this
changed when SARS-CoV was identified, paving the
way for severe forms of the disease in humans (22).
Our previous experience with the outbreaks of other
coronaviruses, like SARS and MERS, suggests that
the mode of transmission in COVID-19 as mainly
human-to-human transmission via direct contact,
droplets, and fomites (25). Recent studies have
demonstrated that the virus could remain viable for
hours in aerosols and up to days on surfaces; thus,
aerosol and fomite contamination could play potent
roles in the transmission of SARS-CoV-2 (257).
The immune response against coronavirus is vital
to control and get rid of the infection. However,
maladjusted immune responses may contribute to the
immunopathology of the disease, resulting in
impairment of pulmonary gas exchange.
Understanding the interaction between CoVs and
host innate immune systems could enlighten our
understanding of the lung inflammation associated
with this infection (24).
SARS is a viral respiratory disease caused by a
formerly unrecognized animal CoV that originated
from the wet markets in southern China after
adapting to the human host, thereby enabling
transmission between humans (90). The SARS
outbreak reported in 2002 to 2003 had 8,098
confirmed cases with 774 total deaths (9.6%) (93).
The outbreak severely affected the Asia Pacific
region, especially mainland China (94). Even though
the case fatality rate (CFR) of SARS-CoV-2
(COVID-19) is lower than that of SARS-CoV, there
exists a severe concern linked to this outbreak due to its epidemiological similarity to influenza viruses
(95, 279). This can fail the public health system,
resulting in a pandemic (96).
MERS is another respiratory disease that was
first reported in Saudi Arabia during the year 2012.
The disease was found to have a CFR of around 35%
(97). The analysis of available data sets suggests that
the incubation period of SARS-CoV-2, SARS-CoV,
and MERS-CoV is in almost the same range. The
longest predicted incubation time of SARS-CoV-2 is
14 days. Hence, suspected individuals are isolated
for 14 days to avoid the risk of further spread (98).
Even though a high similarity has been reported
Even though a high similarity has been reported
between the genome sequence of the new
coronavirus (SARS-CoV-2) and SARS-like CoVs,
the comparative analysis recognized a furin-like
cleavage site in the SARS-CoV-2 S protein that is
missing from other SARS-like COVs (99). The furin-
like cleavage site is expected to play a role in the life
cycle of the virus and disease pathogenicity and
might even act as a therapeutic target for furin
inhibitors. The highly contagious nature of SARS-
CoV-2 compared to that of its predecessors might be
the result of a stabilizing mutation that occurred in
the endosome-associated-protein-like domain of
nsp2 protein.
Similarly, the destabilizing mutation near the
phosphatase domain of nsp3 proteins in SARS-CoV-
2 could indicate a potential mechanism that
differentiates it from other CoVs (100). Even though
the CFR reported for COVID-19 is meager
compared to those of the previous SARS and MERS
outbreaks, it has caused more deaths than SARS and
MERS combined (101). Possibly related to the viral
pathogenesis is the recent finding of an 832
nucleotide (nt) deletion in ORF8, which appears to
reduce the replicative fitness of the virus and leads to
attenuated phenotypes of SARS-CoV-2 (256).
Coronavirus is the most prominent example of a
Coronavirus is the most prominent example of a
virus that has crossed the species barrier twice from
wild animals to humans during SARS and MERS
outbreaks (79, 102). The possibility of crossing the
species barrier for the third time has also been
suspected in the case of SARS-CoV-2 (COVID-19).
Bats are recognized as a possible natural reservoir
host of both SARS-CoV and MERS-CoV infection.
In contrast, the possible intermediary host is the
palm civet for SARS-CoV and the dromedary camel
for MERS-CoV infection (102). Bats are considered
the ancestral hosts for both SARS and MERS (103).
Bats are also considered the reservoir host of human
coronaviruses like HC0V-229E and HCOV-NL63
(104). In the case of COVID-19, there are two
possibilities for primary transmission: it can be
transmitted either through intermediate hosts, similar
to that of SARS and MERS, or directly from bats
(103). The emergence paradigm put forward in the
SARS outbreak suggests that SARS-CoV originated
from bats (reservoir host) and later jumped to civets
(intermediate host) and incorporated changes within
the receptor-binding domain (RBD) to improve
binding to civet ACE2. This civet-adapted virus,
during their subsequent exposure to humans at live
markets, promoted further adaptations that resulted
in the epidemic strain (104). Transmission can also
markets, promoted Turther adaptations that resulted
in the epidemic strain (104). Transmission can also
occur directly from the reservoir host to humans
without RBD adaptations. The bat coronavirus that is
currently in circulation maintains specific “poised”
spike proteins that facilitate human infection without
the requirement of any mutations or adaptations
(105). Altogether, different species of bats carry a
massive number of coronaviruses around the world
(106).
The high plasticity in receptor usage, along with
the feasibility of adaptive mutation and
recombination, may result in frequent interspecies
transmission of coronavirus from bats to animals and
humans (106). The pathogenesis of most bat
coronaviruses is unknown, as most of these viruses
are not isolated and studied (4). Hedgehog
coronavirus HKU31, a Betacoronavirus, has been
identified from amur hedgehogs in China. Studies
show that hedgehogs are the reservoir of
Betacoronavirus, and there is evidence of
recombination (107).
The current scientific evidence available on
MERS infection suggests that the significant
reservoir host, as well as the animal source of MERS
infection in humans, is the dromedary camels (97).
The infected dromedary camels may not show any
visible signs of infection. making it challenging to
visible signs of infection, making it challenging to
identify animals actively excreting MERS-CoV that
has the potential to infect humans. However, they
may shed MERS-CoV through milk, urine, feces,
and nasal and eye discharge and can also be found in
the raw organs (108). In a study conducted to
evaluate the susceptibility of animal species to
MERS-CoV infection, llamas and pigs were found to
be susceptible, indicating the possibility of MERS-
CoV circulation in animal species other than
dromedary camels (109).
Following the outbreak of SARS in China,
SARS-CoV-like viruses were isolated from
Himalayan palm civets (Paguma larvata) and
raccoon dogs (Nyctereutes procyonoides) found in a
live-animal market in Guangdong, China. The
animal isolates obtained from the live-animal market
retained a 29-nucleotide sequence that was not
present in most of the human isolates (78). These
findings were critical in identifying the possibility of
interspecies transmission in SARS-CoV. The higher
diversity and prevalence of bat coronaviruses in this
region compared to those in previous reports indicate
a host/pathogen coevolution. SARS-like
coronaviruses also have been found circulating in the
Chinese horseshoe bat (Rhinolophus sinicus)
populations. The in vitro and in vivo studies carried
populations. The in vitro and in vivo studies carried
out on the isolated virus confirmed that there is a
potential risk for the reemergence of SARS-CoV
infection from the viruses that are currently
circulating in the bat population (105).
CLINICAL PATHOLOGY OF SARS-CoV-2
(COVID-19)
The disease caused by SARS-CoV-2 is also
named severe specific contagious pneumonia
(SSCP), Wuhan pneumonia, and, recently, COVID-
19 (110). Compared to SARS-CoV, SARS-CoV-2
has less severe pathogenesis but has superior
transmission capability, as evidenced by the rapidly
increasing number of COVID-19 cases (111). The
incubation period of SARS-CoV-2 in familial
clusters was found to be 3 to 6 days (112). The mean
incubation period of COVID-19 was found to be 6.4
days, ranging from 2.1 to 11.1 days (113). Among an
early affected group of 425 patients, 59 years was the
median age, of which more males were affected
(114). Similar to SARS and MERS, the severity of
this nCoV is high in age groups above 50 years (2,
115). Symptoms of COVID-19 include fever, cough,
myalgia or fatigue, and, less commonly, headache,
hemoptysis, and diarrhea (116, 282). Compared to
the SARS-CoV-2-infected patients in Wuhan during
the initial stages of the outbreak, only mild
symptoms were noticed in those patients that are
infected by human-to-human transmission (14).
The initial trends suggested that the mortality
associated with COVID-19 was less than that of
previous outbreaks of SARS (101). The updates
obtained from countries like China, Japan, Thailand,
and South Korea indicated that the COVID-19
patients had relatively mild manifestations compared
to those with SARS and MERS (4). Regardless of
the coronavirus type, immune cells, like mast cells,
that are present in the submucosa of the respiratory
tract and nasal cavity are considered the primary
barrier against this virus (92). Advanced in-depth
analysis of the genome has identified 380 amino acid
substitutions between the amino acid sequences of
SARS-CoV-2 and the SARS/SARS-like
coronaviruses. These differences in the amino acid
sequences might have contributed to the difference
in the pathogenic divergence of SARS-CoV-2 (16).
Further research is required to evaluate the possible
differences in tropism, pathogenesis, and
transmission of this novel agent associated with this
change in the amino acid sequence. With the current
outbreak of COVID-19, there is an expectancy of a
significant increase in the number of published
studies about this emerging coronavirus, as occurred
with SARS and MERS (117).
SARS-CoV-2 invades the lung parenchyma,
resulting in severe interstitial inflammation of the
lungs. This is evident on computed tomography (CT)
images as ground-glass opacity in the lungs. This
lesion initially involves a single lobe but later
expands to multiple lung lobes (118). The
histological assessment of lung biopsy samples
obtained from COVID-19-infected patients revealed
diffuse alveolar damage, cellular fibromyxoid
exudates, hyaline membrane formation, and
desquamation of pneumocytes, indicative of acute
respiratory distress syndrome (119). It was also
found that the SARS-CoV-2-infected patients often
have lymphocytopenia with or without leukocyte
abnormalities. The degree of lymphocytopenia gives
an idea about disease prognosis, as it is found to be
positively correlated with disease severity (118).
Pregnant women are considered to have a higher risk
of getting infected by COVID-19. The coronaviruses
can cause adverse outcomes for the fetus, such as
intrauterine growth restriction, spontaneous abortion,
preterm delivery, and perinatal death.
Nevertheless, the possibility of intrauterine
maternal-fetal transmission (vertical transmission) of
CoVs is low and was not seen during either the
SARS- or MERS-CoV outbreak (120). However,
SARS- or MERS-CoV outbreak (120). However,
there has been concern regarding the impact of
SARS-CoV-2/COVID-19 on pregnancy. Researchers
have mentioned the probability of in utero
transmission of novel SARS-CoV-2 from COVID-
19-infected mothers to their neonates in China based
upon the rise in IgM and IgG antibody levels and
cytokine values in the blood obtained from newborn
infants immediately postbirth; however, RT-PCR
failed to confirm the presence of SARS-CoV-2
genetic material in the infants (283). Recent studies
show that at least in some cases, preterm delivery
and its consequences are associated with the virus.
Nonetheless, some cases have raised doubts for the
likelihood of vertical transmission (240–243).
COVID-19 infection was associated with
pneumonia, and some developed acute respiratory
distress syndrome (ARDS). The blood biochemistry
indexes, such as albumin, lactate dehydrogenase, C-
reactive protein, lymphocytes (percent), and
neutrophils (percent) give an idea about the disease
severity in COVID-19 infection (121). During
COVID-19, patients may present leukocytosis,
leukopenia with lymphopenia (244),
hypoalbuminemia, and an increase of lactate
dehydrogenase, aspartate transaminase, alanine
aminotransferase, bilirubin, and, especially, D-dimer
aminotransferase, bilirubin, and, especially, D-dimer
(244). Middle-aged and elderly patients with primary
chronic diseases, especially high blood pressure and
diabetes, were found to be more susceptible to
respiratory failure and therefore, had poorer
prognoses. Providing respiratory support at early
stages improved the disease prognosis and facilitated
recovery (18). The ARDS in COVID-19 is due to the
occurrence of cytokine storms that results in
exaggerated immune response, immune regulatory
network imbalance, and, finally, multiple-organ
failure (122). In addition to the exaggerated
inflammatory response seen in patients with
COVID-19 pneumonia, the bile duct epithelial cell-
derived hepatocytes upregulate ACE2 expression in
liver tissue by compensatory proliferation that might
result in hepatic tissue injury (123).
CORONAVIRUSES IN ANIMALS AND
ZOONOTIC LINKS-A BRIEF
VIEWPOINT
Coronavirus can cause disease in several species
of domestic and wild animals, as well as humans
(23). The different animal species that are infected
with CoV include horses, camels, cattle, swine, dogs,
cats, rodents, birds, ferrets, minks, bats, rabbits,
snakes, and various other wild animals (20, 30, 79,
snakes, and various other wild animals (20, 30, 79,
93, 124, 125, 287). Coronavirus infection is linked to
different kinds of clinical manifestations, varying
from enteritis in cows and pigs, upper respiratory
disease in chickens, and fatal respiratory infections
in humans (30).
Among the CoV genera, Alphacoronavirus and
Betacoronavirus infect mammals, while
Gammacoronavirus and Deltacoronavirus mainly
infect birds, fishes, and, sometimes, mammals (27,
29, 106). Several novel coronaviruses that come
under the genus Deltacoronavirus have been
discovered in the past from birds, like Wigeon
coronavirus HKU20, Bulbul coronavirus HKU11,
Munia coronavirus HKU13, white-eye coronavirus
HKU16, night-heron coronavirus HKU19, and
common moorhen coronavirus HKU21, aswell as
from pigs (porcine coronavirus HKU15) (6, 29).
Transmissible gastroenteritis virus (TGEV), porcine
epidemic diarrhea virus (PEDV), and porcine
hemagglutinating encephalomyelitis virus (PHEV)
are some of the coronaviruses of swine. Among
them, TGEV and PEDV are responsible for causing
severe gastroenteritis in young piglets with
noteworthy morbidity and mortality. Infection with
PHEV also causes enteric infection but can cause
encephalitis due to its ability to infect the nervous
system (30).
Bovine coronaviruses (BoCoVs) are known to
infect several domestic and wild ruminants (126).
BoCoV inflicts neonatal calf diarrhea in adult cattle,
leading to bloody diarrhea (winter dysentery) and
respiratory disease complex (shipping fever) in cattle
of all age groups (126). BoCoV-like viruses have
been noted in humans, suggesting its zoonotic
potential as well (127). Feline enteric and feline
infectious peritonitis (FIP) viruses are the two major
feline CoVs (128), where feline CoVs can affect the
gastrointestinal tract, abdominal cavity (peritonitis),
respiratory tract, and central nervous system (128).
Canines are also affected by CoVs that fall under
different genera, namely, canine enteric coronavirus
in Alphacoronavirus and canine respiratory
coronavirus in Betacoronavirus, affecting the enteric
and respiratory tract, respectively (129, 130). IBV,
under Gammacoronavirus, causes diseases of
respiratory, urinary, and reproductive systems, with
substantial economic losses in chickens (131, 132).
In small laboratory animals, mouse hepatitis virus,
rat sialodacryoadenitis coronavirus, and guinea pig
and rabbit coronaviruses are the major CoVs
associated with disease manifestations like enteritis,
hepatitis, and respiratory infections (10, 133).
Swine acute diarrhea syndrome coronavirus
system (30).
Bovine coronaviruses (BoCoVs) are known to
infect several domestic and wild ruminants (126).
BoCoV inflicts neonatal calf diarrhea in adult cattle,
leading to bloody diarrhea (winter dysentery) and
respiratory disease complex (shipping fever) in cattle
of all age groups (126). BoCoV-like viruses have
been noted in humans, suggesting its zoonotic
potential as well (127). Feline enteric and feline
infectious peritonitis (FIP) viruses are the two major
feline CoVs (128), where feline CoVs can affect the
gastrointestinal tract, abdominal cavity (peritonitis),
respiratory tract, and central nervous system (128).
Canines are also affected by CoVs that fall under
different genera, namely, canine enteric coronavirus
in Alphacoronavirus and canine respiratory
coronavirus in Betacoronavirus, affecting the enteric
and respiratory tract, respectively (129, 130). IBV,
under Gammacoronavirus, causes diseases of
respiratory, urinary, and reproductive systems, with
substantial economic losses in chickens (131, 132).
In small laboratory animals, mouse hepatitis virus,
rat sialodacryoadenitis coronavirus, and guinea pig
and rabbit coronaviruses are the major CoVs
associated with disease manifestations like enteritis,
hepatitis, and respiratory infections (10, 133).
Swine acute diarrhea syndrome coronavirus
Swine acute diarrhea syndrome coronavirus
(SADS-CoV) was first identified in suckling piglets
having severe enteritis and belongs to the genus
Alphacoronavirus (106). The outbreak was
associated with considerable scale mortality of
piglets (24,693 deaths) across four farms in China
(134). The virus isolated from the piglets was almost
identical to and had 95% genomic similarity with
horseshoe bat (Rhinolophus species) coronavirus
HKU2, suggesting a bat origin of the pig virus (106,
134, 135). It is also imperative to note that the
SADS-CoV outbreak started in Guangdong province,
near the location of the SARS pandemic origin
(134). Before this outbreak, pigs were not known to
be infected with bat-origin coronaviruses. This
indicates that the bat-origin coronavirus jumped to
pig by breaking the species barrier. The next step of
this jump might not end well, since pigs are
considered the mixing vessel for influenza A viruses
due to their ability to be infected by both human and
avian influenza A viruses (136).
Similarly, they may act as the mixing vessel for
coronaviruses, since they are in frequent contact with
both humans and multiple wildlife species.
Additionally, pigs are also found to be susceptible to
infection with human SARS-CoV and MERS-CoV,
making this scenario a nightmare (109, 137). It is
only a matter of time before another zoonotic
coronavirus results in an epidemic by jumping the
so-called species barrier (287).
The host spectrum of coronavirus increased when
a novel coronavirus, namely, SW1, was recognized
in the liver tissue of a captive beluga whale
(Delphinapterus leucas) (138). In recent decades,
several novel coronaviruses were identified from
different animal species. Bats can harbor these
viruses without manifesting any clinical disease but
are persistently infected (30). They are the only
mammals with the capacity for self-powered flight,
which enables them to migrate long distances, unlike
land mammals. Bats are distributed worldwide and
also account for about a fifth of all mammalian
species (6). This makes them the ideal reservoir host
for many viral agents and also the source of novel
coronaviruses that have yet to be identified. It has
become a necessity to study the diversity of
coronavirus in the bat population to prevent future
outbreaks that could jeopardize livestock and public
health. The repeated outbreaks caused by bat-origin
coronaviruses calls for the development of efficient
molecular surveillance strategies for studying
Betacoronavirus among animals (12), especially in
the Rhinolophus bat family (86). Chinese bats have
high commercial value, since they are used in
high commercial value, since they are used in
traditional Chinese medicine (TCM). Therefore, the
handling of bats for trading purposes poses a
considerable risk of transmitting zoonotic CoV
epidemics (139)
Due to the possible role played by farm and wild
animals in SARS-CoV-2 infection, the WHO, in
their novel coronavirus (COVID-19) situation report,
recommended the avoidance of unprotected contact
with both farm and wild animals (25). The live-
animal markets, like the one in Guangdong, China,
provides a setting for animal coronaviruses to
amplify and to be transmitted to new hosts, like
humans (78). Such markets can be considered a
critical place for the origin of novel zoonotic
diseases and have enormous public health
significance in the event of an outbreak. Bats are the
reservoirs for several viruses; hence, the role of bats
in the present outbreak cannot be ruled out (140). In
a qualitative study conducted for evaluating the
zoonotic risk factors among rural communities of
southern China, the frequent human-animal
interactions along with the low levels of
environmental biosecurity were identified as
significant risks for the emergence of zoonotic
disease in local communities (141, 142).
The comprehensive sequence analysis of the
The comprehensive sequence analysis of the
SARS-CoV-2 RNA genome identified that the CoV
from Wuhan is a recombinant virus of the bat
coronavirus and another coronavirus of unknown
origin. The recombination was found to have
happened within the viral spike glycoprotein, which
recognizes the cell surface receptor. Further analysis
of the genome based on codon usage identified the
snake as the most probable animal reservoir of
SARS-CoV-2 (143). Contrary to these findings,
another genome analysis proposed that the genome
of SARS-CoV-2 is 96% identical to bat coronavirus,
reflecting its origin from bats (63). The involvement
of bat-derived materials in causing the current
outbreak cannot be ruled out. High risk is involved
in the production of bat-derived materials for TCM
practices involving the handling of wild bats. The
use of bats for TCM practices will remain a severe
risk for the occurrence of zoonotic coronavirus
epidemics in the future (139).
Furthermore, the pangolins are an endangered
species of animals that harbor a wide variety of
viruses, including coronaviruses (144). The
coronavirus isolated from Malayan pangolins (Manis
javanica) showed a very high amino acid identity
with COVID-19 at E (100%), M (98.2%), N
(96.7%), and S genes (90.4%). The RBD of S protein
(96.7%), and S genes (90.4%). The RBD of S protein
in CoV isolated from pangolin was almost identical
(one amino acid difference) to that of SARS-CoV-2.
A comparison of the genomes suggests
recombination between pangolin-CoV-like viruses
with the bat-CoV-RaTG13-like virus. All this
suggests the potential of pangolins to act as the
intermediate host of SARS-CoV-2 (145).
Human-wildlife interactions, which are
increasing in the context of climate change (142), are
further considered high risk and responsible for the
emergence of SARS-CoV. COVID-19 is also
suspected of having a similar mode of origin. Hence,
to prevent the occurrence of another zoonotic
spillover (1), exhaustive coordinated efforts are
needed to identify the high-risk pathogens harbored
by wild animal populations, conducting surveillance
among the people who are susceptible to zoonotic
spillover events (12), and to improve the biosecurity
measures associated with the wildlife trade (146).
The serological surveillance studies conducted in
people living in proximity to bat caves had earlier
identified the serological confirmation of SARS-
related CoVs in humans. People living at the
wildlife-human interface, mainly in rural China, are
regularly exposed to SARS-related CoVs (147).
These findings will not have any significance until a
These findings will not have any significance until a
significant outbreak occurs due to a virus-like
SARS-CoV-2.
There is a steady increase in the reports of
COVID-19 in companion and wild animals around
the world. Further studies are required to evaluate
the potential of animals (especially companion
animals) to serve as an efficient reservoir host that
can further alter the dynamics of human-to-human
transmission (330). To date, two pet dogs (Hong
Kong) and four pet cats (one each from Belgium and
Hong Kong, two from the United States) have tested
positive for SARS-CoV-2 (335). The World
Organization for Animal Health (OIE) has confirmed
the diagnosis of COVID-19 in both dogs and cats
due to human-to-animal transmission (331). The
similarity observed in the gene sequence of SARS-
CoV-2 from an infected pet owner and his dog
further confirms the occurrence of human-to-animal
transmission (333). Even though asymptomatic,
feline species should be considered a potential
transmission route from animals to humans (326).
However, currently, there are no reports of SARS-
CoV-2 transmission from felines to human beings.
Based on the current evidence, we can conclude that
cats are susceptible to SARS-CoV-2 and can get
infected by human beings. However, evidence of cat
infected by human beings. However, evidence of cat-
to-human transmission is lacking and requires
further studies (332). Rather than waiting for firmer
evidence on animal-to-human transmission,
necessary preventive measures are advised, as well
as following social distancing practices among
companion animals of different households (331).
One of the leading veterinary diagnostic companies,
IDEXX, has conducted large-scale testing for
COVID-19 in specimens collected from dogs and
cats. However, none of the tests turned out to be
positive (334).
In a study conducted to investigate the potential
of different animal species to act as the intermediate
host of SARS-CoV-2, it was found that both ferrets
and cats can be infected via experimental inoculation
of the virus. In addition, infected cats efficiently
transmitted the disease to naive cats (329). SARS-
CoV-2 infection and subsequent transmission in
ferrets were found to recapitulate the clinical aspects
of COVID-19 in humans. The infected ferrets also
shed virus via multiple routes, such as saliva, nasal
washes, feces, and urine, postinfection, making them
an ideal animal model for studying disease
transmission (337). Experimental inoculation was
also done in other animal species and found that the
dogs have low susceptibility, while the chickens,


মন্তব্যসমূহ
একটি মন্তব্য পোস্ট করুন